| Review Article |
|
|
National Research Laboratory for Cardiac Stem Cell, Seoul National University College of Medicine, Seoul, Korea
Corresponding Author: hyo-Soo Kim, Department of Internal Medi-tural and catalytic protein in epithelial cells cine, Seoul national University hospital, 28 Yongon-dong, chongro-gu, Seoul 110-744, South Korea. Tel: 82-2-2072-2226; Fax: 82-2-766-8904; ILK was first discovered at 1996 by Hannigan and E-mail: hyosoo@snu.ac.kr.
| |
ABSTRACT |
|
|
INTRODUCTION |
|
|
CONCLUSION |
|
|
ACKNOWLEDGEMENT |
|
|
CONFLICT OF INTEREST STATEMENT |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Integrin-linked kinase (ILK) is an intracellular molecule that binds
to the cytoplasmic domain of β1 and β3-integrin. It has been previously
demonstrated in various epithelial cell lines to mediate the ‘outside-in’
signals into the cells and to control the survival of these cells by controlling
the phosphorylation of various downstream proteins, such as protein kinase B/Akt
(PKB/Akt). We now present in this review the important role of ILK in the
vascular system with particular emphasis on its role in endothelial cells (ECs).
The results presented here demonstrate that ILK is essential for the proper
function, structure and survival of ECs and finally, for the process of
neovascularization.
KEY WORDS:
integrin-linked kinase; vascular biology; blood vessel; endothelium
|
|
INTRODUCTION |
|---|
Coronary artery disease and cerebrovascular disease are devastating status of acute vessel occlusion in diseased vessels that are already narrowed enough by atheroscle- rotic process. According to a Korean data, this disease ac-count for more than 50% of the total cause of death and and the burden continues increase. These diseases are caused basically by ischemic damage of the target organ, for ex- ample acute myocardial infarction from coronary artery and ischemic stroke from intracerebral artery occlusion. The prevention of these cardiovascular diseases involves the meticulous use of multiple drugs such as aspirin and beta-blockers, but these drugs are not enough to regenerate the organ or tissue that has been already damaged by preceding vascular events. People are now focused on tissue regeneration (1) and one method to do so is the supply and growth of new vessels into the ischemic tissue. It is now well-known that blood vessels are not only a ‘passive conduit’ for passage of blood components and gas, but an ‘active participant’ of several physiological/pathological processes that we encounter in everyday life in the clinics. Integrin-linked kinase (ILK) is an intracellular molecule that has been under intense investigation nowadays. Several groups, including our laboratory, have focused on its role in the vascular system in the past couple of years, and have demonstrated that the role of ILK is not only important in tumorigenesis in several epithelial cells but that it is also a critical molecule mediating the survival and function of endothelial cells (ECs). It has also been demonstrated that not only is ILK important in ECs but it is also critical in the function (2) and recruitment of endothelial progenitor cells (EPCs) to the appropriate tissue (3). Therefore ILK provides a new, interesting target of genetic modulation in either enhancing or suppressing the process of neovascularization. Unraveling the natural role of ILK in the vascular system is expected to be a cornerstone in this field. In this review, we will introduce the role of ILK in the vascular system, together with the results of the work in our laboratory, which has lead to some interesting results concerning the role of ILK in EPCs as well.
Integrin-linked Kinase: Its discovery as a both structural and catalytic protein in epithelial cells
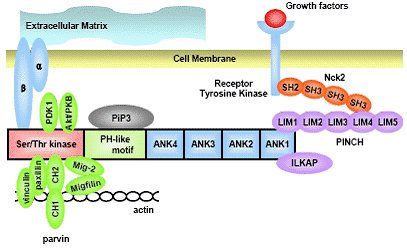
ILK was first discovered at 1996 by Hannigan and Dedhar as an intracellular molecule that binds to the cy-toplasmic domain of β1 and β3-integrin and is expressed ubiquitously in various tissues and cells (4). It is mainly comprised of 3 structurally different regions (5). There are 4 ankyrin (ANK) repeats in its N-terminus and then a pleckstrin homology (PH)-like motif. Lastly, there exists a domain at the C-terminus which shares significant homol-ogy with other protein catalytic domains. The concern on ILK was initially focused on its ability to mediate vari-ous protein-to-protein interactions and also its ability to transduce the external mechanical stimuli to an internal chemical stimulus.
Up to date, there are 10 proteins that can bind to ILK and these can be largely divided into 3 parts (5). The list of these proteins is listed in Table 1. The first and second sets of proteins are adaptor proteins and integrins that connect the extracellular matrix with the intracellular structural proteins such as actin cytoskeletons. Particularly INter-esting Cys-His rich motif (PINCH), a focal adhesion pro-tein, and the ANK1 domain of ILK binds to each other to maintain the cell shape (6, 7). PINCH is comprised of 5 LIM domains (8) and it has been shown previously that it can undergo considerable conformational change upon binding of ILK (9). In addition, PINCH binds to various receptor tyrosine kinase via Nck2 (10). In vivo, PINCH is required for the assembly of muscle-dense bodies in stud-ies of Caenorhabditis elegans (C. elegans) (11). Disrup-tion of ILK in Drosophila lead to the detachment of actin filaments from the muscle attachment site, suggesting that the PINCH-ILK interaction is indeed important for main-taining the structure of the cells in vivo (12).
|
Parvins, that consists of 3 set of proteins, α-parvin/ actopaxin/calponin homology ILK binding protein (CH-ILKBP), β-parvin/affixin and γ-parvin, is a protein that binds to the kinase domain of ILK (13-16) and acts as a critical protein maintaining the cell shape and mechano- transduction into the cell (17). α-parvin/actopaxin/CH-ILKBP interacts directly with actin, as well as indirectly by binding with paxillin (13, 15). β-parvin can also link ILK with actin by interacting with α-actinin (18). The proteins that interact with γ-parvin are not identified yet.However, there has been interesting, arguing results on the role of γ-parvin in hematopoietic cells. Although the purpose and the viewpoints of the experiments were very different, deletion of γ-parvin in vitro has been shown to result in defective monocyte migration and spreading to ECM (19), whereas disruption of γ-parvin in vivo lead to no gross abnormality in the development of hematopoietic system (20).
Mitogen inducible gene-2(Mig-2),a mammalian homo-log of UNC-112 in C. elegans, is also a molecule that inter-acts with the filaminin component of actin (21). Mig-2 has also been shown to be a candidate molecule linking actin to ILK (22). In an experimental model of others, ILK was required for the proper localization of structural proteins to integrin foci, suggesting that ILK is a critical protein mediating outside-in and inside-out signals in cells (22).
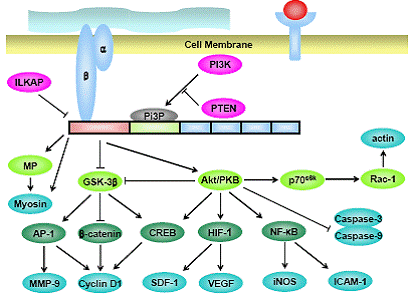
The third set of molecules are catalytic proteins, such as PKB/Akt (23) and ILK associated protein (ILKAP) (24). The kinase activity of ILK is under regulation of certain proteins such as ILKAP (24) and phosphoinositide-3-ki-nase (PI3-kinase) (25) and this peculiar kinase activity of ILK needs PH motif, as mutants of the PH-domain in ILK leads to the inability of ILK to phosphorylate Akt/PKB (26). Several proteins that directly interact with ILK are demonstrated in Figure 1.
|
Together with these structural functions of ILK, the catalytic function of this protein has also been under in-tense investigation until recently. The role of ILK in vari-ous epithelial cells has been shown by analyzing the ILK activity in either growth factor or extracellular matrix (ECM) stimulated cells, or in ILK-overexpressing cells. Overexpression of ILK in various epithelial cell lines has been shown to activate PKB/Akt (26), mitogen activated protein kinase (MAP kinase) (27) and inactivate glyco-gen synthase kinase 3β (GSK-3β) (25, 28). Moreover it has also been shown to downregulate E-cadherin (29, 30) and activate the nuclear translocation of β-catenin (30), pos-sibly by inactivating GSK-3β. Cyclin D1 expression has also been shown to be upregulated by activation of acti-vating protein -1 (AP-1) through inactivation of GSK-3β (28, 31). But these trafficking of intracellular proteins are not always consistent, as the activation/inactivation of Akt, GSK-3β and β-catenin does not take place in Dro-sophila system after ILK overexpression (32), suggesting that the downstream molecules in one type of cells are not so in another.
One of the function that has been of particular inter-est in ILK is its activity to suppress apoptosis and anoikis (suspension-induced apoptosis) (23, 33). These effects are mainly due to the suppression or activation of proteins that are involved in cell survival and apoptosis such as PKB/ Akt (25) and caspase-3 and 8 (33). It has been demonstrat-ed in mammary epithelial cell lines that overexpression of ILK leads to the suppression of anoikis in a PKB/Akt-de-pendent manner (33) and these phenomenon has also been shown to occur in various cancer cell lines such as breast cancer (27) and prostate cancer (23). The overall substrates of ILK are depicted in Figure 2.
|
Taken together, these findings indicate ILK as a unique protein that can act as both structural and catalytic protein in epithelial cells. These findings have been adopted and utilized in the field of oncology and ILK holds as a prom-ising target of many neoplastic disease such as prostate cancer and breast cancer (34).
The discovery of ILK in the vascular system
ILK has been under intense investigation in various cell lines in epithelial lineages but the role of ILK in the vascular system has been under investigation very recent-ly. This part of the paper would focus on the role of ILK in vascular biology with particular emphasis on its role in ECs and the process of neovascularization.
1) The role of ILK in endothelial cells. The possibil- ity of the role of ILK in ECs, has first been known in 2001 by Zhang et.al., who demonstrated that oxidized LDL (ox-LDL) treatment in human umbilical vein endothelial cells (HUVECs) resulted in the transcriptional and trans-lational upregulation of ILK (35). The overexpression of ILK lead to the suppression of ox-LDL-induced apoptosis and this phenomenon was mediated by various intracel-lular molecules involved in apoptosis and survival, such as p38 MAP kinase, bcl-2, bcl-xL, caspase-3 and 9. These findings shed light into the possibility that the pathways found in epithelial cell lineages would also happen in ECs and that genetic enhancement of ILK in ECs would be protective. This possibility was partially confirmed by Gonzalez-Santiago et.al. shortly afterwards, who showed that the HUVECs cultured in collagen type I, a pathologic condition in which HUVECs are not ordinarily exposed to, showed decreased endothelial nitric oxide synthase (eNOS) activity and also, decreased ILK kinase activity (36). This phenomenon was rescued by overexpression of ILK in collagen type I cultured HUVECs, thus confirm-ing that ILK could act as a molecule in reducing various stress. The ability of ILK to rescue HUVECs under spe-cific stress, such as oxidative stress and abnormal ECM, was further expanded to conditions such as starvation (nu-trient deprivation) and cell suspension (anchorage depriva-tion) in another paper from our group (2). The authors took the advantage that ILK was transiently upregulated after nutrient and anchorage deprivation in HUVECs and that enhancing the natural increase of ILK would naturally lead to the suppression of apoptosis under these stressful conditions. These effects of ILK was partially dependent on several intracellular molecules, such as PKB/Akt and GSK-3β, previously described to be downstream of ILK in various epithelial cells, supporting that many of the intra-cellular pathways found in the mammalian epithelial cells would also occur in ECs as well.
Another part of the role of ILK in ECs mainly stem from the idea that ILK is upstream of a well-known intra-cellular molecule PKB/Akt, which in turn is known to be important in the function and survival of ECs in vitro and in vivo. Also, vascular endothelial growth factor (VEGF), a strong mitogen and survival factor in ECs, is known to be secreted in various cancer cells such as prostate can-cer (37) and breast cancer (38, 39), which in turn activates PKB/Akt in ECs (40, 41). Therefore it is logical to think that ILK would be important in the intracellular signaling and the function in ECs. In accordance to this hypothesis, the paper by Tan and Dedhar shows exactly the critical im-portance of ILK in ECs. According to this paper, prostate cancer cells secrete VEGF in a ILK-dependent manner via pathways involving Akt 劤 劤m-TOR (mammalian target of rapamycin) 劤HIF-1 (hypoxia inducible factor -1) (42). Also, the functions of ECs by the secreted VEGF were as-sessed by survival, migration and invasion assay and also by tube formation and EC sprouting assay. This action of VEGF in ECs was completely abrogated by blockade of ILK with transfection of si-RNA of ILK, confirming that the above functions of VEGF in ECs were ILK-dependent. The aforementioned action of VEGF via ILK was again confirmed by the papers from Kaneko and Basaki, pub-lished shortly afterwards. ILK was found to be bound to VEGFR-2 in response to VEGF stimulation and the main functions of VEGF was suppressed by KD-ILK (kinase deficient form of ILK) transfection (43). These phenomena emphasize the kinase function of ILK in maintaining the basic function of ECs and that ILK is a critical molecule mediating the VEGF 劤 PKB/Akt pathway.
However, it was shortly followed by another paper by Vouret-Craviari and van Obberghen-Schilling, who dem-onstrated that ILK is important in EC adhesion and capil-lary formation (44). This role of ILK was associated with distribution of α5β1 integrin on the cell surface and also with fibrillar fibronectin matrix. Interestingly, silencing ILK with RNA interference was shown to induce mor-phological changes in the structure of ECs, changing the shape of EC to a rather round shape. Also, silencing of ILK induced the cellular expression of certain integrins, such as α5β1 and αvβ3 integrin, and finally, increased cell adhesion to ECM, whereas cell spreading, migration and tube formation was impaired in ILK-depleted cells. In ac-cordance with these findings, Watanabe and Basaki also noticed that transfection of KD-ILK plasmid resulted in decreased tube formation in HUVECs stimulated with VEGF (45). Therefore, one can implicate from the above findings that ILK is indispensable in the dynamic features of ECs, such as proliferation, migration and tube forma-tion and that the kinase function of ILK is more important in these features than its structural association with other intracellular proteins.
The natural role of ILK in EC was further implicated to in vivo findings by other groups which lead to some very interesting findings. Friedrich and Gerszten were among the first to find that conditional knock-out of ILK in ECs leads to a defect in embryogenesis by placental insufficiency caused via decreased labyrinthine vascu-larization in vivo and massive apoptosis of ECs in vitro (46). But discordant to the previous findings by Vouret- Craviari and van Obberghen-Schilling, deletion of ILK in ECs resulted in a characteristic detachment of ECs from the matrix and loss of actin fibers. Whether the discrep-ancy between these two independent experiments is due to the experimental conditions or the technique used to knock-out ILK (si-RNA vs. Cre-recombinase conditional knock-out) remains to be confirmed. The finding in this paper is concordant with another previous paper, in which ILK knock-out resulted in a lethal phenotype, mainly by a defect in actin rearrangement but also defective vascular development (47).
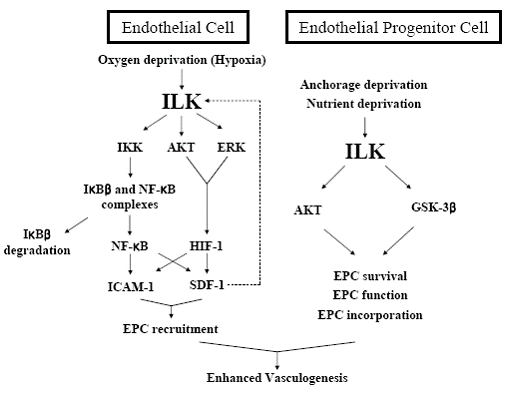
In addition to the previous works to reveal the basic role - especially the structural function - of ILK in ECs, efforts to control the process of vasculogenesis by genetic modulation of ILK has also lead to some interesting, new findings mainly from our laboratory. The previous find-ings in our laboratory that ILK is transiently elevated in various stress conditions in ECs also held true in EPCs from adult human peripheral blood (2) and this phenom-enon has given the idea that enhancing ILK would also reinforce the survival pathway involving PKB/Akt, GSK-3β and finally vasculogenesis. In vitro, ILK-overexpress-ing EPCs showed superior ability in incorporating into the EC layer and this was successfully implicated to the in vivo model. ILK-overexpressing EPCs were success- ful in salvaging ischemic tissue even with only 5% of the cells given in the control group. This finding has the major ability to overcome the obstacle in the sparcity of EPCs in adult human peripheral blood by genetic modification. Together with the findings that ILK has a critical role in EPC function, it was also demonstrated in the following paper from our laboratory that ILK was also critical in EPC recruitment to the appropriate ischemic tissue. ILK was naturally enhanced in its amount and also in its ki-nase activity in hypoxic ECs. The abrogation of the kinase activity of ILK in hypoxic ECs lead to a significantly sup-pressed expression of stromal cell derived factor -1 (SDF-1) and intercellular adhesion molecule -1 (ICAM-1), the two key molecules found to be important in EPC recruit-ment to ischemic tissue (48, 49). Finally, genetic engineer-ing of ILK in hypoxic tissue lead to the control of EPC recruitment in ischemic tissue and finally vasculogenesis. These findings of the important role of ILK in postnatal vasculogenesis can be summarized into Figure 3.
|
ILK has also been found to have a potential role in EC dysfunction. Although there is limited data on this subject, it has been demonstrated rather earlier that ILK is involved in EC-ECM interaction and endothelial dys-function. Seeding EC on collagen I rather than a normal ECM, collagen IV, decreased the expression of eNOS and also the production of nitric oxide from ECs, the effect of which was reversed by overexpression of ILK (36). The aforementioned control of eNOS by ILK was not further investigated in depth but one can guess that both the ki- nase activity (activation of PKB/Akt by ILK, which in turn can activated eNOS (50, 51)) and the structural function (maintenance of actin cytoskeleton by ILK, which in turn regulates eNOS activity (52)) of ILK can be involved in this process. This finding indicates that ILK can be an-other new, biologic marker of endothelial dysfunction. To-gether with the previous data, it has also been shown that culture of EC in collagen I leads to decreased activity of ILK and also increased expression of MCP-1 and adhe-sion of peripheral blood monocytes to ECs (53). This effect was abrogated by transfection of ILK into EC, which also suggests that ILK is involved in endothelial dysfunction. Whether these in vitro findings have further implication into in vivo findings, such as atherosclerotic disease, also remains to be confirmed.
2) The Role of ILK in Platelets. Although there is a limited amount of data in this field, the role of ILK in platelets has also been demonstrated in studies done in platelet activation. The potentiality of the involvement of ILK in this process is a fancy target of research, concern- ing that ILK is a intracellular protein that binds to β1 and β3-integrin, the two key surface molecules involved in in-flammation and platelet activation. Although the experi-mental methods and design differ slightly from each other, a few groups have commonly demonstrated that the kinase activity of ILK increases after stimulation of platelets (54- 56). Moreover, ILK has been shown in platelets to mediate the ‘activation’ signal into the cell - for example, activation of platelets by adenosine diphosphate or thrombin - and in turn, to ‘outside’ activity -for example, increased phos-phorylation of β-integrin -. These effects has been con-firmed and further implicated into the adhesion activity of platelets (57). Platelets activated by thrombin exhibited in-creased kinase activity of ILK and ultimately, to increased adhesion to collagen. Therefore, the role of ILK may be further implicated to in vivo models of acute thrombosis and this remains to be further investigated.
3) The Role of ILK in Vascular Smooth Muscle Cells. In contrast to the well-investigated role of ILK in ECs, its role in vascular smooth muscle cells (VSMCs) is largely not understood. Specifically, there has recently been one journal reporting that the amount and activity of ILK is specifically upregulated by angiotensin-II and downregu-lated by statin treatment in vitro and in vivo (58). This paper implicates that ILK may play a critical role in ath-erosclerosis and that it would be a good potential target of anti-atherosclerotic management. As in the role of ILK in ECs, the investigation of its role in VSMCs is further war-ranted in the future.
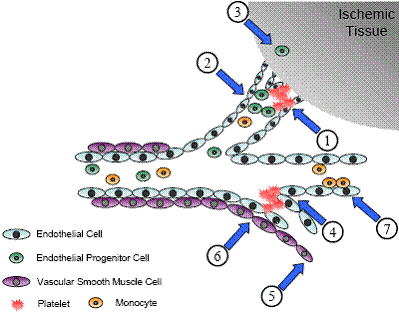
Taken together, the role of ILK in the vascular system that has been verified and also, its role that needs further study, can be summarized into Figure 4.
|
| |
|---|
ILK is an intracellular protein that is ubiquitously ex-pressed in various types of cells and tissues. As its role in several neoplastic diseases is now being revealed, it is also known to be a key molecule also in the vascular bi-ology, particularly ECs. But, it remains to be further in- vestigated whether it has any role in other components of the blood vessel, such as vascular smooth muscle cells. In addition, many of the findings of ILK in this field have been focused on in vitro findings but its role in various in vivo models is very limited. Together with the forecoming results of ILK in various human disease models, we hope to gain an integrated understanding of the role of ILK in vascular biology.
|
|
ACKNOWLEDGEMENT |
|---|
This study was supported by grants from the program of National Research Laboratory for Cardiovascular Stem Cell, KOSEF, Republic of Korea.
|
|
CONFLICT OF INTEREST STATEMENT |
|---|
The authors declare that no conflict of interest exists.
|
|
REFERENCES
|
|---|
1. Laflamme MA, Murry CE. Regenerating the heart. Nat Biotechnol. 2005;23:845-56.
2. Cho HJ, Youn SW, Cheon SI, Kim TY, Hur J, Zhang SY, Lee SP, Park KW, Lee MM, Choi YS, Park YB, Kim HS. Regulation of endothelial cell and endothelial progenitor cell survival and vasculogenesis by integrin-linked kinase. Arterioscler Thromb Vasc Biol. 2005;25:1154- 60.
3. Lee SP, Youn SW, Cho HJ, Li L, Kim TY, Yook HS, Chung JW, Hur J, Yoon CH, Park KW, Oh BH, Park YB, Kim HS. Integrin-linked kinase, a hypoxia-responsive molecule, controls postnatal vasculogenesis by recruitment of endothelial progenitor cells to ischemic tissue. Circulation. 2006;114:150-9.
4. Hannigan GE, Leung-Hagesteijn C, Fitz-Gibbon L, Coppolino MG, Radeva G, Filmus J, Bell JC, Dedhar S. Regulation of cell adhesion and anchorage-dependent growth by a new beta 1-integrin-linked protein kinase. Nature. 1996;379:91-6.
5. Wu C, Dedhar S. Integrin-linked kinase (ILK) and its interactors: a new paradigm for the coupling of extracellular matrix to actin cytoskeleton and signaling complexes. J Cell Biol. 2001;155:505-10.
6. Tu Y, Li F, Goicoechea S, Wu C. The LIM-only protein PINCH directly interacts with integrin-linked kinase and is recruited to integrin-rich sites in spreading cells. Mol Cell Biol. 1999;19:2425-34.
7. Li F, Zhang Y, Wu C. Integrin-linked kinase is localized to cell-matrix focal adhesions but not cell-cell adhesion sites and the focal adhesion localization of integrin-linked kinase is regulated by the PINCH-binding ANK repeats. J Cell Sci. 1999;112 ( Pt 24):4589-99.
8. Wu C. Integrin-linked kinase and PINCH: partners in regulation of cell-extracellular matrix interaction and signal transduction. J Cell Sci. 1999;112 ( Pt 24):4485-9.
9. Wu C. Integrin-linked kinase and PINCH: partners in regulation of cell-extracellular matrix interaction and signal transduction. J Cell Sci. 1999;112 ( Pt 24):4485-9.
10. Vaynberg J, Fukuda T, Chen K, Vinogradova O, Velyvis A, Tu Y, Ng L, Wu C, Qin J. Structure of an ultraweak protein-protein complex and its crucial role in regulation of cell morphology and motility. Mol Cell. 2005;17:513-23.
11. Hobert O, Moerman DG, Clark KA, Beckerle MC, Ruvkun G. A conserved LIM protein that affects muscular adherens junction integrity and mechanosensory function in Caenorhabditis elegans. J Cell Biol. 1999;144:45-57.
12. Clark KA, McGrail M, Beckerle MC. Analysis of PINCH function in Drosophila demonstrates its requirement in integrin-dependent cellular processes. Development. 2003;130:2611-21.
13. Tu Y, Huang Y, Zhang Y, Hua Y, Wu C. A new focal adhesion protein that interacts with integrin-linked kinase and regulates cell adhesion and spreading. J Cell Biol. 2001;153:585-98.
14. Yamaji S, Suzuki A, Sugiyama Y, Koide Y, Yoshida M, Kanamori H, Mohri H, Ohno S, Ishigatsubo Y. A novel integrin-linked kinasebinding protein, affixin, is involved in the early stage of cell-substrate interaction. J Cell Biol. 2001;153:1251-64.
15. Nikolopoulos SN, Turner CE. Actopaxin, a new focal adhesion protein that binds paxillin LD motifs and actin and regulates cell adhesion. J Cell Biol. 2000;151:1435-48.
16. Olski TM, Noegel AA, Korenbaum E. Parvin, a 42 kDa focal adhesion protein, related to the alpha-actinin superfamily. J Cell Sci. 2001;114:525-38.
17. Lin X, Qadota H, Moerman DG, Williams BD. C. elegans PAT-6/ actopaxin plays a critical role in the assembly of integrin adhesion complexes in vivo. Curr Biol. 2003;13:922-32.
18. Yamaji S, Suzuki A, Kanamori H, Mishima W, Yoshimi R, Takasaki H, Takabayashi M, Fujimaki K, Fujisawa S, Ohno S, Ishigatsubo Y. Affixin interacts with alpha-actinin and mediates integrin signaling for reorganization of F-actin induced by initial cell-substrate interaction. J Cell Biol. 2004;165:539-51.
19. Yoshimi R, Yamaji S, Suzuki A, Mishima W, Okamura M, Obana T, Matsuda C, Miwa Y, Ohno S, Ishigatsubo Y. The gamma-parvin-integrin- linked kinase complex is critically involved in leukocyte-substrate interaction. J Immunol. 2006;176:3611-24.
20. Chu H, Thievessen I, Sixt M, Lammermann T, Waisman A, Braun A, Noegel AA, Fassler R. gamma-Parvin is dispensable for hematopoiesis, leukocyte trafficking, and T-cell-dependent antibody response. Mol Cell Biol. 2006;26:1817-25.
21. Tu Y, Wu S, Shi X, Chen K, Wu C. Migfilin and Mig-2 link focal adhesions to filamin and the actin cytoskeleton and function in cell shape modulation. Cell. 2003;113:37-47.
22. Mackinnon AC, Qadota H, Norman KR, Moerman DG, Williams BD. C. elegans PAT-4/ILK functions as an adaptor protein within integrin adhesion complexes. Curr Biol. 2002;12:787-97.
23. Persad S, Attwell S, Gray V, Delcommenne M, Troussard A, Sanghera J, Dedhar S. Inhibition of integrin-linked kinase (ILK) suppresses activation of protein kinase B/Akt and induces cell cycle arrest and apoptosis of PTEN-mutant prostate cancer cells. Proc Natl Acad Sci U S A. 2000;97:3207-12.
24. Leung-Hagesteijn C, Mahendra A, Naruszewicz I, Hannigan GE. Modulation of integrin signal transduction by ILKAP, a protein phosphatase 2C associating with the integrin-linked kinase, ILK1. Embo J. 2001;20:2160-70.
25. Delcommenne M, Tan C, Gray V, Rue L, Woodgett J, Dedhar S. Phosphoinositide- 3-OH kinase-dependent regulation of glycogen synthase kinase 3 and protein kinase B/AKT by the integrin-linked kinase. Proc Natl Acad Sci U S A. 1998;95:11211-6.
26. Persad S, Attwell S, Gray V, Mawji N, Deng JT, Leung D, Yan J, Sanghera J, Walsh MP, Dedhar S. Regulation of protein kinase B/Akt-serine 473 phosphorylation by integrin-linked kinase: critical roles for kinase activity and amino acids arginine 211 and serine 343. J Biol Chem. 2001;276:27462-9.
27. White DE, Cardiff RD, Dedhar S, Muller WJ. Mammary epithelialspecific expression of the integrin-linked kinase (ILK) results in the induction of mammary gland hyperplasias and tumors in transgenic mice. Oncogene. 2001;20:7064-72.
28. D’Amico M, Hulit J, Amanatullah DF, Zafonte BT, Albanese C, Bouzahzah B, Fu M, Augenlicht LH, Donehower LA, Takemaru K, Moon RT, Davis R, Lisanti MP, Shtutman M, Zhurinsky J, Ben-Ze’ev A, Troussard AA, Dedhar S, Pestell RG. The integrin-linked kinase regulates the cyclin D1 gene through glycogen synthase kinase 3beta and cAMP-responsive element-binding protein-dependent pathways. J Biol Chem. 2000;275:32649-57.
29. Somasiri A, Howarth A, Goswami D, Dedhar S, Roskelley CD. Overexpression of the integrin-linked kinase mesenchymally transforms mammary epithelial cells. J Cell Sci. 2001;114:1125-36.
30. Novak A, Hsu SC, Leung-Hagesteijn C, Radeva G, Papkoff J, Montesano R, Roskelley C, Grosschedl R, Dedhar S. Cell adhesion and the integrin-linked kinase regulate the LEF-1 and beta-catenin signaling pathways. Proc Natl Acad Sci U S A. 1998;95:4374-9.
31. Dedhar S. Cell-substrate interactions and signaling through ILK. Curr Opin Cell Biol. 2000;12:250-6.
32. Zervas CG, Gregory SL, Brown NH. Drosophila integrin-linked kinase is required at sites of integrin adhesion to link the cytoskeleton to the plasma membrane. J Cell Biol. 2001;152:1007-18.
33. Attwell S, Roskelley C, Dedhar S. The integrin-linked kinase (ILK) suppresses anoikis. Oncogene. 2000;19:3811-5.
34. Hannigan G, Troussard AA, Dedhar S. Integrin-linked kinase: a cancer therapeutic target unique among its ILK. Nat Rev Cancer. 2005;5:51- 63.
35. Zhang X, Hu K, Li CY. Protection against oxidized low-density lipoprotein- induced vascular endothelial cell death by integrin-linked kinase. Circulation. 2001;104:2762-6.
36. Gonzalez-Santiago L, Lopez-Ongil S, Rodriguez-Puyol M, Rodriguez- Puyol D. Decreased nitric oxide synthesis in human endothelial cells cultured on type I collagen. Circ Res. 2002;90:539-45.
37. Doll JA, Reiher FK, Crawford SE, Pins MR, Campbell SC, Bouck NP. Thrombospondin-1, vascular endothelial growth factor and fibroblast growth factor-2 are key functional regulators of angiogenesis in the prostate. Prostate. 2001;49:293-305.
38. Linderholm B, Tavelin B, Grankvist K, Henriksson R. Vascular endothelial growth factor is of high prognostic value in node-negative breast carcinoma. J Clin Oncol. 1998;16:3121-8.
39. Yoshiji H, Gomez DE, Shibuya M, Thorgeirsson UP. Expression of vascular endothelial growth factor, its receptor, and other angiogenic factors in human breast cancer. Cancer Res. 1996;56:2013-6.
40. Gerber HP, McMurtrey A, Kowalski J, Yan M, Keyt BA, Dixit V, Ferrara N. Vascular endothelial growth factor regulates endothelial cell survival through the phosphatidylinositol 3’-kinase/Akt signal transduction pathway. Requirement for Flk-1/KDR activation. J Biol Chem. 1998;273:30336-43.
41. Fujio Y, Walsh K. Akt mediates cytoprotection of endothelial cells by vascular endothelial growth factor in an anchorage-dependent manner. J Biol Chem. 1999;274:16349-54.
42. Tan C, Cruet-Hennequart S, Troussard A, Fazli L, Costello P, Sutton K, Wheeler J, Gleave M, Sanghera J, Dedhar S. Regulation of tumor angiogenesis by integrin-linked kinase (ILK). Cancer Cell. 2004;5:79- 90.
43. Kaneko Y, Kitazato K, Basaki Y. Integrin-linked kinase regulates vascular morphogenesis induced by vascular endothelial growth factor. J Cell Sci. 2004;117:407-15.
44. Vouret-Craviari V, Boulter E, Grall D, Matthews C, Van Obberghen- Schilling E. ILK is required for the assembly of matrix-forming adhesions and capillary morphogenesis in endothelial cells. J Cell Sci. 2004;117:4559-69.
45. Watanabe M, Fujioka-Kaneko Y, Kobayashi H, Kiniwa M, Kuwano M, Basaki Y. Involvement of integrin-linked kinase in capillary/tube-like network formation of human vascular endothelial cells. Biol Proced Online. 2005;7:41-7.
46. Friedrich EB, Liu E, Sinha S, Cook S, Milstone DS, MacRae CA, Mariotti M, Kuhlencordt PJ, Force T, Rosenzweig A, St-Arnaud R, Dedhar S, Gerszten RE. Integrin-linked kinase regulates endothelial cell survival and vascular development. Mol Cell Biol. 2004;24:8134-44.
47. Sakai T, Li S, Docheva D, Grashoff C, Sakai K, Kostka G, Braun A, Pfeifer A, Yurchenco PD, Fassler R. Integrin-linked kinase (ILK) is required for polarizing the epiblast, cell adhesion, and controlling actin accumulation. Genes Dev. 2003;17:926-40.
48. Ceradini DJ, Kulkarni AR, Callaghan MJ, Tepper OM, Bastidas N, Kleinman ME, Capla JM, Galiano RD, Levine JP, Gurtner GC. Progenitor cell trafficking is regulated by hypoxic gradients through HIF- 1 induction of SDF-1. Nat Med. 2004;10:858-64.
49. Yoon CH, Hur J, Oh IY, Park KW, Kim TY, Shin JH, Kim JH, Lee CS, Chung JK, Park YB, Kim HS. Intercellular adhesion molecule-1 is upregulated in ischemic muscle, which mediates trafficking of endothelial progenitor cells. Arterioscler Thromb Vasc Biol. 2006;26:1066- 72.
50. Fulton D, Gratton JP, McCabe TJ, Fontana J, Fujio Y, Walsh K, Franke TF, Papapetropoulos A, Sessa WC. Regulation of endotheliumderived nitric oxide production by the protein kinase Akt. Nature. 1999;399:597-601.
51. Dimmeler S, Fleming I, Fisslthaler B, Hermann C, Busse R, Zeiher AM. Activation of nitric oxide synthase in endothelial cells by Aktdependent phosphorylation. Nature. 1999;399:601-5.
52. Laufs U, Endres M, Stagliano N, Amin-Hanjani S, Chui DS, Yang SX, Simoncini T, Yamada M, Rabkin E, Allen PG, Huang PL, Bohm M, Schoen FJ, Moskowitz MA, Liao JK. Neuroprotection mediated by changes in the endothelial actin cytoskeleton. J Clin Invest. 2000;106:15-24.
53. Ruiz-Torres MP, G PER-R, Rodriguez-Puyol M, Rodriguez-Puyol D, Diez-Marqu Eacute SM. The Leukocyte-Endothelial Cell Interactions are Modulated by Extracellular Matrix Proteins. Cell Physiol Biochem. 2006;17:221-32.
54. Barry FA, Gibbins JM. Protein kinase B is regulated in platelets by the collagen receptor glycoprotein VI. J Biol Chem. 2002;277:12874-8.
55. Pasquet JM, Noury M, Nurden AT. Evidence that the platelet integrin alphaIIb beta3 is regulated by the integrin-linked kinase, ILK, in a PI3-kinase dependent pathway. Thromb Haemost. 2002;88:115-22.
56. Yamaji S, Suzuki A, Kanamori H, Mishima W, Takabayashi M, Fujimaki K, Tomita N, Fujisawa S, Ohno S, Ishigatsubo Y. Possible role of ILK-affixin complex in integrin-cytoskeleton linkage during platelet aggregation. Biochem Biophys Res Commun. 2002;297:1324-31.
57. Stevens JM, Jordan PA, Sage T, Gibbins JM. The regulation of integrin- linked kinase in human platelets: evidence for involvement in the regulation of integrin alpha 2 beta 1. J Thromb Haemost. 2004;2:1443- 52.
58. Friedrich EB, Clever YP, Wassmann S, Werner N, Bohm M, Nickenig G. Role of integrin-linked kinase in vascular smooth muscle cells: regulation by statins and angiotensin II. Biochem Biophys Res Commun. 2006;349:883-9.