| ORIGINAL ARTICLE |
|
|
1 Post Graduate Department of Biochemistry and Biotechnology, Cell and Molecular Therapeutics Laboratory, Oriental Institute of Science and Technology, Vidyasagar University, Midnapore-721102, West Bengal, India;
2 Founder and Secretary, Agricure Biotech Research Society, Epidemiology and Human Health Division, Midnapore-721101, India
Corresponding Author: Dr. Smarajit Maiti, Reader and Head, Post Graduate Department of Biochemistry and Biotechnology, Cell & Molecular Therapeutics Lab, OIST, Vidyasagar University, Midnapore-721102, India. Tel: 9474504269; E-mail: maitism@rediffmail.com.
Running title: Antioxidative pharmacological efficacies of Bellamya bengalensis against arsenic-toxicity
| |
ABSTRACT |
| INTRODUCTION | |
|
|
MATERIALS AND METHODS |
|
|
RESULTS |
|
|
DISCUSSION |
|
|
CONCLUSIONS |
|
|
CONFLICTING INTERESTS |
|
|
ACKNOWLEDGEMENT |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Chronic arsenic exposure results in cancer. Some therapeutic agents show inadequate-potency/ side-effects in arsenic-toxicity treatment. The Bellamya bengalensis, an edible snail has long been used by rural people comprised of both ethnic and nonethnic groups as traditional medicine in several health-anomalies/ liver-disorders. In an attempt to investigate the possible protective and therapeutic effect against arsenic induced rat tissue damage are conferred by antioxidative mechanism and attenuation of pro-inflammatory response, the extract of B. bengalensis was tested in arsenic intoxicated rat model. Here, Bellamya bengalensis flesh-extract (BBE, 1 g/kg bw/day for 28days) was tested concomitantly in arsenic-intoxicated (0.6 ppm/kg bw/day for 28days) rat, in in-vitro rat liver slices (in Krebs-ringer buffer for 2 and 4 hours treatment with sodium arsenite alone or with BBE). In the rat, BBE strongly prevented arsenic-induced oxidative/necrotic damages to the intestinal epithelial tissue and liver-tissue/DNA by strengthening the antioxidant-system as shown in Non-protein soluble thiol (NPSH), Superoxide Dismutase(SOD) & catalase results which are clearly reflected in DNA-ladder/comet-assay/histo-architecture results. Arsenic alone decreased catalase and SOD activities in-vivo and in-vitro (H2O2/arsenite redox-stress to dialyzed-concentrated SOD) and also decreased antioxidative signaling molecules i.e. NPSH, serum nitric-oxide (NO) levels. At the same time, arsenic increased the tissue malondialdehyde resulting in DNA-breakage/liver-damage which except NO, were restrained by BBE that constitutes high-level of phosphorus/ascorbate/free-thiols. Moreover, an arsenic-induced increase in pro-inflammatory cytokine TNF-α was restored terminating an acute-phase-reaction. This study, for the first-time, shows the efficiencies of some organism/animal extract in hepatic and intestinal tissue challenged with a high level of arsenic with comparison to the natural level water contamination in West Bengal, India. Our present outcome may be utilized for the development of some protective/therapeutic component against arsenic toxicity from this aquatic organism. Further studies are necessary for more conclusive comments.
KEY WORDS: Arsenic toxicity; Tissue necrosis; DNA damage; Carcinogenesis; Ethnomedicinal Effects|
|
INTRODUCTION |
|---|
Arsenic is a naturally occurring component that is present ubiquitously in the environment. Humans can be manifested to arsenic through food and in some regions, through drinking water (1). The most affected areas in the world are Bangladesh and West Bengal, India. The World Health Organization amended the maximum permissible limit of arsenic to be 50 μg/l (2). Epidemiological studies have shown that chronic exposure to inorganic arsenic is associated with cancers of the skin, lung, and bladder (3). In chronic arsenic ingestion, arsenic accumulates in the liver, kidneys, heart, and lungs and smaller amounts in the gastrointestinal tract, and spleen (4). Arsenic is metabolized by reduction and methylation reactions, catalyzed by glutathione-S-transferase omega-1 (GSTO1) and arsenic (III) methyltransferase (AS3MT) (5). The gastrointestinal (GI) tract presents itself as the first organ susceptible to attack by ingested xenobiotics (6). The major site of absorption is the small intestine by an electrogenic process involving a proton (H+) gradient (7). The optimal pH for arsenic absorption is 5.0 (8). The environment of the bowel is almost neutral (pH is approximately 7.0) due to pancreatic bicarbonate secretion (9). Many freshwater mollusks are important food source for humans such as snails like Bellamya bengalensis. For a long time these snails/ aquatic organisms have ethnopharmacologically importance (10). B. bengalensis, a highly protein rich organism is extensively consumed as food in the rural part of India, Bangladesh and other countries (11-13). This organism is extensively used for disturbance of eyesight, vitamins deficiencies and helps in controlling diarrhea and gastric disorders (14).
In relation to arsenic therapeutics, recently, some herbal and phytochemicals remediation of arsenic-induced tissue toxicity has been shown (15-17) but none has a conclusive and satisfactory level of efficacy. So, in the current study, the Bellamya bengalensis extract has been tested to screen its therapeutic effects in arsenic-induced gastrointestinal toxicity in in vitro/ in vivo rat model.
|
|
MATERIALS AND METHODS |
|---|
Bellamya bengalensis Extract (BBE) Preparation
Mature Bellamya sp. were collected from local ponds at Midnapore region and carefully washed to remove clay particle then blotted to remove excess moisture by blotting paper. 20 gm of flesh (edible part) of Bellamya bengalensis was added to 50 ml distilled water and homogenized by electric homogenizer carefully to prepare 40% BBE. Then the total homogenate tissue was collected in cold condition and centrifuged by cold centrifuge machine at 4°C temperature for 10 mints at 10,000 rpm. The supernatant was collected and used freshly or stored for a couple of days at –4°C.
Animal Selection and Treatment
Female albino rats weighing 150-160 gm. were acclimatized for 10 days at 12-h light-dark cycle, 32 ± 2°C temperature, 50–70% humidity in the institutional animal resource facility. Those were fed a standard pellet diet (Hindustan Lever, Mumbai, India) and water ad libitum. Studies were carried out in accordance with the National Institutes of Health, USA guidelines and the institutional ethical concerns (no. 3.i ec2014) were maintained throughout the investigation. Rats were randomly distributed into three groups having six in each. Animals of group-II and group-III are fed with 0.5 ml drinking water containing sodium arsenite at a concentration of 0.6 ppm (600 µg/L) /100 g body weight / day for 28 days. Initially, several dose-response studies of arsenic were conducted on a rat model. The present dose range usually does not cause animal mortality but exposure for a moderate time period (≥ 3 weeks) increased the level of intestinal toxicity and other clinical marker suggesting a significant level of cellular toxicity (16).
Supplementation with BBE to Arsenic-Exposed Rats
The group-I designated as control is supplied with the same amount of drinking water for stipulated duration. Group-III is supplemented with BB extract by gavages at a concentration of 100 mg tissue of Bellamya bengalensis /100 g body weight/ day for 28 days. On the day 29, animals are exposed to light anesthesia (by ether), blood is collected using a disposable syringe (21-gauge needle) and serum is separated and organs required for biochemical and histological examinations were dissected out.
In the in situ experimentation, rat liver slices were incubated with arsenite or different oxidant/reductant alone or with BBE or BBV in Kreb’s buffer. Post centrifuged (12,000 × g) cytosolic fraction was prepared from tissue homogenate and that was cleaned by dialysis membrane (Spectrum Lab, USA) and concentrated with the Amicon centrifugal filter units (Millipore, USA, 6-8kd MWCO) to negate small molecules interferences. The concentrated fraction was tested for SOD1 activity by NBT test in polyacrylamide gel as described in the previous section.
Determination of Catalase Activities
Catalase activity is assayed by a colorimetric method (18). Dichromate in acetic acid is converted to perchromic acid and then to chromic acetate when heated in the presence of H2O2. The chromic acetate formed is measured at 620 nm. The catalase preparation is allowed to split H2O2 for different periods of time. The reaction is stopped at different time intervals by the addition of a dichromate-acetic acid mixture and the remaining H2O2 is determined as chromic acetate. One unit of the enzymatic activity is expressed as a mole of H2O2 consumed per min per mg protein.
Estimation of Malondialdehyde (MDA) and Conjugated diene Level
Tissues were homogenized in 0.1 M chilled phosphate buffer (pH 7.4). The MDA assay is conducted following the protocol of (19) with a slight modification. The reaction was conducted in the cold room with buffer as possibly deprived with O2 to minimize ROS formation during experimental procedure. To chelate iron and reduce its interference in peroxidation reaction of unsaturated fatty acid, 1 mM EDTA is used in the reaction mixture. To reduce the interference caused by a yellow-orange color produced by some carbohydrates, the reaction mixture is heated at 80°C instead of 100°C. Finally, the MDA is measured and calculated using the molar extinction coefficient of MDA (1.56 × 105 cm2/mmol) (20).
CDs are determined by a standard method (21). In brief, lipids are extracted with chloroform– methanol (2:1), followed by centrifugation at 3000 rpm for 5 min, lipid residues are then dissolved in 1.5 mL of cyclohexane and the absorbance is measured at 233 nm to determine the amount of hydroperoxide formed.
Assay of Superoxide Dismutase (SOD)
A tablet of nitro blue tetrazolium (NBT) was dissolved in 30 ml water and the nondenaturing (10%) acrylamide gel (Protean-II, Bio-Rad electrophoresis system, USA) was soaked with it for 30 min with shaking. The gel was then shaken in 40 ml SOD1 (cytosolic SOD, also known as Cu-Zn SOD) solution (0.028 M tetramethylethylenediamine (TEMED), 2.8 × 10-5 M riboflavin, and 0.036 M potassium phosphate at pH 7.8) for 15 min. The soaked gel was placed on a clean acetate sheet and illuminated for 5 to 15 min. The gel became purple except at the position containing SOD1 (22) The gel was scanned when the maximum contrast between the band and background has been achieved.
Estimation of Non-protein Soluble Thiol
The Nonprotein soluble thiol (NPSH) in intestinal cells homogenate (prepared in 0.1 M phosphate buffer, pH 7.4) is determined by standard 5, 50-dithiobis-2-nitrobenzoic acid (DTNB) method with a slight modification (23). The experimental was conducted in cold room with some protease inhibitor in the buffer solution. This minimizes the endogenous protein degradation during the experimental procedure. In brief, the protein is precipitated by trichloroacetic acid and clear cytosol is added to 0.1 M sodium phosphate buffer containing 5 mM DTNB. The level of NPSH is determined against a glutathione (GSH) standard curve.
Evaluation of General Toxicity: Liver and Kidney Function Tests
Serum glutamate oxaloacetate transaminase (SGOT), glutamate pyruvate transaminase (SGPT), alkaline phosphatase (ALP), urea, creatinine, and uric acid are measured from the rat’s serum of different groups by standard protocol with the assay kits (Ranbaxy, India or other reputed company).
Analysis of Hematopoietic Profile
Total count and differential count (DC) were done manually by using a Spencer Bright-Line hemacytometer blood test kit (American Optical, Scientific Instrumental Division, Buffalo, NY).
Comet Assay
Cell suspension is prepared after it is scrapped and collected by a Teflon scrapper from the intestinal parts of animals after sacrifice and immediately before the assay procedure. It is performed in a cold room and under dimmed light to avoid any possible exposure to UV light. The chilled 1X PBS [calcium and magnesium free and Dimethyl sulfoxide (DMSO) added] is used to minimize endoglucanase activities and minimize the iron of hem (from hemoglobin) associated free radical interference. Using blunt-ended tips or thick syringe, cells are prepared homogeneous suspension and clump free. After the cell count cell number is adjusted to 105 cells/mL and kept in microcentrifuge. The alkaline Comet assay is done according to the method of Singh et al. with some minor modifications (24). A total of 75 μL of low melting point agarose (0.5%) in PBS at 37°C is added to a 25 μL cell suspension. The mixture is then dropped onto a microscope slide pre-coated with 1% agarose and a cover slip is placed on it. Once the agarose solidifies the cover slips are removed and the slides are immersed in ice-cold lysis buffer (2.5 mM NaCl, 85 mM EDTA, 10 mM Trizma base, 1% Triton X-100, 10% DMSO, and 1% sodium lauryl sarcosinate, adjusted to pH 10) for 1 h at 4°C. After lysis the slides are washed three times in PBS at room temperature. Slides are then placed in a submarine gel electrophoresis chamber filled with alkaline electrophoresis buffer (0.3 M NaOH and 1 mM EDTA) for 25 min. Then the electrophoresis is done for 30 min at 25 V and the current is adjusted to 300 mA by raising the buffer level. Slides are then neutralized with PBS and stained with a solution of 2 mg/mL ethidium bromide for 3-4 min. Excess stain is removed by washing in water. Slides are read using a fluorescence microscope.
Histology and DNA Fragmentation Analysis
Intestine tissue was embedded in paraffin, serially sectioned at 5 mM, stained with eosin and hematoxylin (Harris) and PAS stain observed under a microscope (Nikon, Eclipse LV100, magnification × 50 ×) to study the histoarchitecture. Intestinal epithelial cells were used for DNA preparation, tissue was treated with 500 mL of lysis buffer (50 mM Tris pH 8.0, 20 mM EDTA, 10 mM NaCl, 1% SDS, 0.5 mg/mL proteinase K) for 20 min on ice (4°C) and centrifuged in cold at 12,000 g for 30 min. The supernatant was extracted for DNA isolation with standardized method (25).
Estimation of Serum Nitric Oxide (NO) Level
Nitric oxide was assayed by use of the methemoglobin method, following a procedure described elsewhere, by use of a Beckman spectrophotometer (model DU6). Assay validity was confirmed by use of an independent chemiluminescence method (26).
Estimation of Serum TNF-α Level
Serum TNF-α level was quantified by ELISA kit (catalog no. MBS355371, MyBioSource, Inc., USA) using monoclonal antibody according to the method (27).
In vitro Experiment in Rat Liver Slices and Intestinal Tissue
Liver slices from albino female rat were incubated under five different experimental condition in Krebs ringer buffer for 2 and 4 hours accordingly, group -1 control (1A, 1B, 1C), group -2 sodium arsenite (NaAsO2) (1 mM) + H2O2 (100 mM) (2A, 2B, 2C), group -3 NaAsO2 (1 mM) (3A, 3B, 3C), group -4 NaAsO2 (1 mM) +H2O2 (100 mM) +100 ul BBE (40%) (4A, 4B, 4C), group -5 NaAsO2 (1 mM) +100 ul BBE (40%) (5A, 5B, 5C). Post centrifuged (12,000 × g) cytosolic fraction was prepared from tissue homogenate and that was cleaned by dialysis membrane (Spectrum Lab, USA) and concentrated with the Amicon centrifugal filter units (Millipore, USA, 6-8kd MWCO) to negate small molecules interferences.
In vitro regulation of SOD activity
The SOD activity was tested by NBT test in polyacrylamide gel as described earlier (22).
Catalase activity assay by gel-zymography
Catalase activity assay was done by gel-zymography using 8% native gel. Chemicals used 0.003% H2O2 (30% solution vol./vol.), 2% ferric chloride (wt./vol.), 2% potassium ferricyanide (wt./vol.). After staining, it has developed a green-blue color with the white band where the enzyme is present. Following separation of the native protein, the catalase enzyme removes the peroxides from the area of the gel it occupies. Removal of peroxide does not allow for the potassium ferricyanide (a yellow substance) to be reduced to potassium ferrocyanide that reacts with ferric chloride to form a Prussian blue precipitate (28).
Estimation of malondialdehyde (MDA) level
The MDA assay was conducted following the protocol as describe earlier (19-21) with a slight modification.
Estimation of non-protein soluble thiol
The NPSH assay was conducted following the protocol as describe earlier (23).
Comet assay in liver slice
The alkaline comet assay was conducted for the confirmation of DNA damage following the guidelines (24) with slight modifications.
In vitro study of Catalase and SOD (Cu-Zn SOD or SOD1) activity after sodium arsenite treatment with free phosphorus and cysteine
Liver slices from albino female rat were incubated under five different experimental condition in Krebs-ringer buffer for 2 or 4 hours accordingly, and its protection by Na2HPO4 and L-cysteine were verified with their concomitant treatment with arsenic. Groups are generated as follows. Group1-control, 2- NaAsO2 (250 µM), 3 -NaAsO2 (250 µM) + H2O2 (100 mM), 4- NaAsO2 (250 µM) + Na2HPO4, 5- NaAsO2 (250 µM) + L- Cysteine (100 mM), 6- NaAsO2 (250 µM) +H2O2 (100 mM) + Na2HPO4, 7- NaAsO2 (250 µM) +H2O2 (100 mM) + L- Cysteine (100 mM). Post centrifuged (12,000 × g) cytosolic fraction was prepared from tissue homogenate prepared after the scheduled incubation period. Those were cleaned by dialysis membrane (Spectrum Lab, USA) and concentrated with the Amicon centrifugal filter units (Millipore, USA, 6-8kd MWCO) to negate small molecules interferences.
Catalase and SOD (Cu-Zn SOD or SOD1) activity by gel-zymography were conducted following the protocol as describe earlier (28, 22).
|
|
RESULTS |
|---|
Status of Oxidative Stress Markers
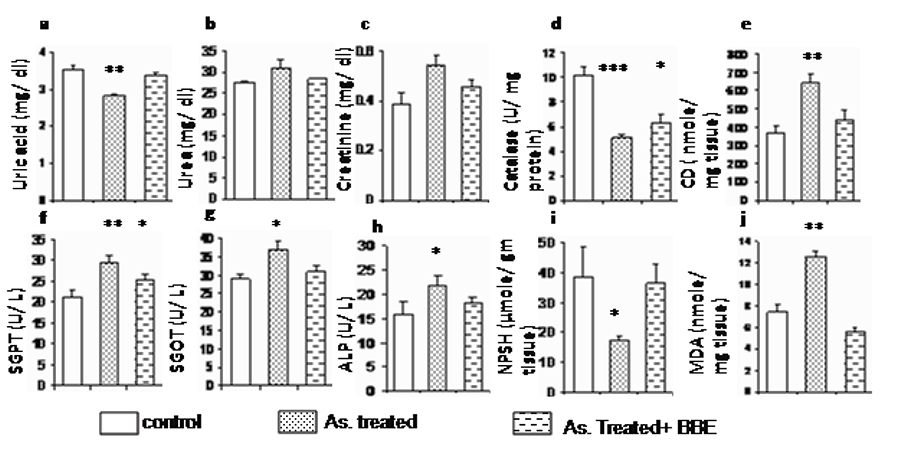
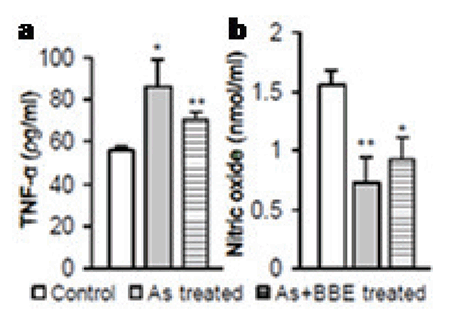
In this set of experimentations, we examined the effects of arsenic on the intestinal tissue antioxidant status. The MDA and CD content in intestinal epithelial cell homogenates significantly increased in the sodium arsenite-exposed rats. However, administration with BBE combined with arsenic prevented MDA and CD elevation when compared to the arsenic only treated group (P<0.001) (Fig. 1e, 1j). There was a significant decrease in intestinal SOD and catalase activities in arsenic-treated rats when compared to control group (P<0.001). Restoration of SOD (Fig. 4c) and catalase (Fig. 1d) activities were observed in the BBE supplementation group. However, administration of BBE in addition to arsenic significantly prevented their elevations (P<0.001) and also regained NPSH level than that of arsenite-exposed rats (Fig. 1i). Arsenic caused a significant increase (p<0.05) in the level of proinflammatory cytokines TNF-α and significant decrease (p<0.01) in NO level in the arsenic-treated groups. Though not significant, appreciable reversal was noticed in both parameters of BBE supplemented group (Fig. 2a, 2b).
General Toxicity: Liver and Kidney Function Test
The result suggests that the serum SGPT, SGOT, and ALP enzymatic activities are significantly increased (p<0.05) in arsenic intoxicated rat suggesting necrotic tissue damage which has been ceased and restored by the BBE supplementation (Fig. 1f, 1g, 1h). Similarly, the increase in the kidney function marker urea and creatinine, restored by the BBE supplemented group in arsenic exposed rats (Fig. 1b, 1c). A significant depletion of serum uric acid by arsenic (P<0.05) and its restoration by BBE are noticed (Fig. 1a).
Hematopoietic Profile
Rats’ hematopoietic profiles (total count and differential count of WBC) are presented in Table 1, these data suggest that increase of total count of WBC in arsenic intoxicated rat blood compared with control group and that was restrained by BBE supplemented group. Results of DC indicate that neutrophil increased by 9% and lymphocyte decreased 8% in arsenic-treated rats when compared to control group and its restoration by BBE are noticed Table 1 and Table 2.
Hepatic Tissue Architecture, DNA Fragmentation and Comet Assay Result
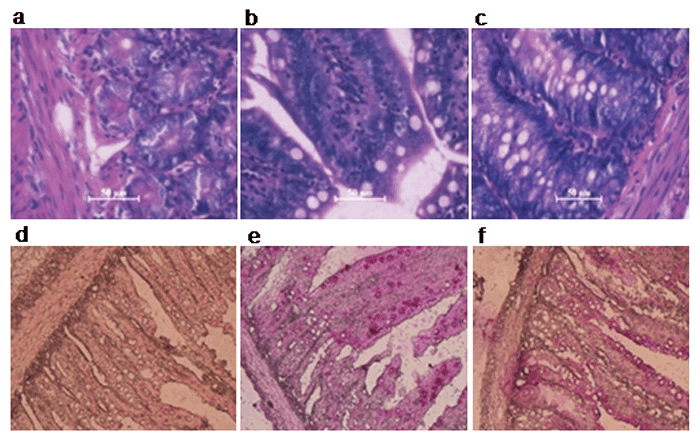
Arsenic ingestion resulted in intestinal tissue disarrangement with lobular/epithelial degeneration. The histopathological evaluations suggest the event of hemorrhages and infiltration of mononuclear cells in the intestine. The layers of muscularis mucosae (inner and outer layer) of intestinal wall are found to be impaired in the arsenic-exposed rat. But the co-administration of BBE shows partial but significant protection which is evident from the picture (Fig. 7a, 7b, 7c). PAS staining suggests the micronecrotic lesions in the tissues (pointed by the arrow head) and impairment in the mucin-producing abilities. It has been significantly restored by the BBE supplementation (Fig. 7d, 7e, 7f).
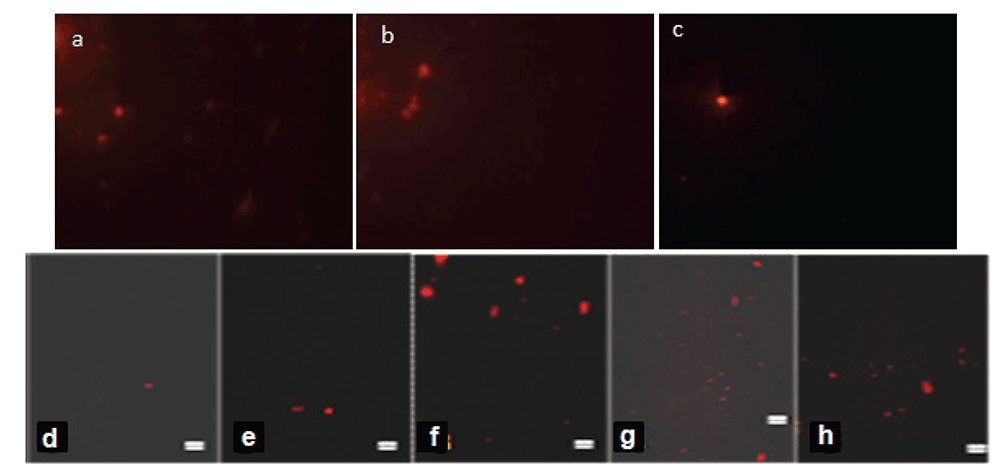
The single cell DNA status (comet assay) basically supports the DNA fragmentation results. The number of comets forming cells is found to be higher in arsenic exposed rat intestinal epithelial cell whereas, in BBE supplemented group the genetic materials of the cells are noticed to be highly protected (Fig. 5a, 5b, 5c).
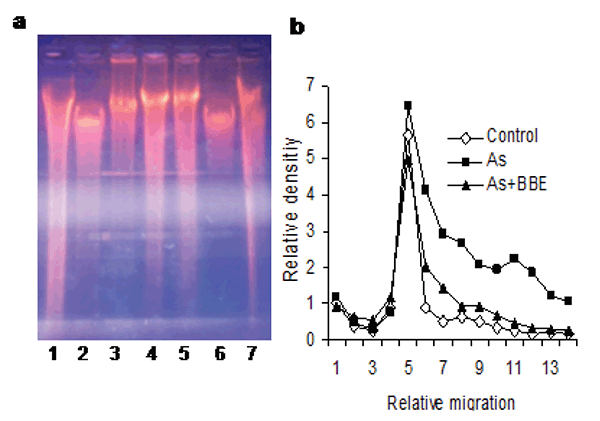
The DNA ladder assay (Fig. 6a). Arsenic-induced DNA damage (laddering in lane 3, 4, 5 in Fig. 6a) is shown to be protected in the lane 6, 7. The band density analysis is shown in Figure 6b.
In vitro experimental data
Comet assay.The comet assay showed that the intestinal DNA in the arsenic-treated cells have damaged as per the (Fig. 5d, 5e) where cells seems to be larger due to the dispersion of DNA as compared to the control (Fig. 5a). Whereas prevention of the said toxicity was observed in the As + BBE (Fig. 5g, 5h).
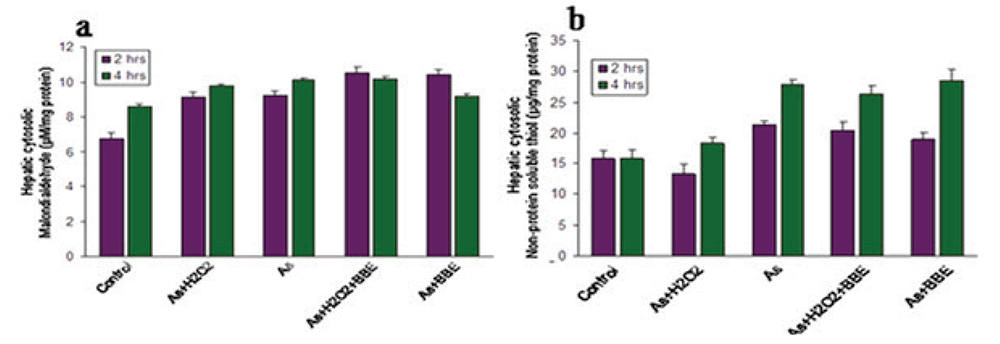
MDA (malondialdehyde) Assay.In the lipid peroxidation study, we notice a moderate increase of lipid peroxidation in liver tissue. In this condition, the BBE treated seems to be more protective of long duration. Whereas, arsenic treatment for a long duration manifested its toxicity in the form of increased lipid-peroxidation and it is possibly circumvented by the higher level of thiol as noticed in NPSH assay. So in can be suggested the lipid peroxidation might have some limited scope for the manifestation in tissue toxicity (Fig. 3a).
NPSH (Nonprotein soluble thiol) Assay.The results from NPSH assay shows that it in increased in liver of As-treated groups in a time dependent manner. But it is significantly increased only in As treated group and all BBE treated group. This suggests that in the case of As toxicity thiol plays an adaptive role and in BBE treated this increase showed an adaptive and protective role against As induce hepatic toxicity. NPSH is the thiol precursor for all other thiol containing biomolecule. Which plays a direct and indirect antioxidant against oxidative stressed induce threatening apart from that and disulfide maintain intercellular redox balance and thiol is the active component of several enzyme receptors or signaling molecule. So thiol-induced protection was noticed in BBE treated group (Fig. 3b).
Superoxide Dismutase in Gel-zymography.Superoxide dismutase (SOD) assay in gel shows that the SOD activity was decreased by the As & H2O2 and As in the lane (3, 4) & (5, 6) respectively than the control (lane 1 & 2). Whereas the SOD activity was regained when the Bellameya extract was used as remediation against the As + H2O2 & As in the lane (7 & 8) and (9 & 10) (Fig. 4b).
Catalase activity in Gel-zymography.Catalase assay in the gel shows that the catalase activity is decreased by the As + H2O2 & As in the lane (3, 4) (5, 6) respectively than the control (lane 1 & 2). The activity of the catalase was regained when the Bellamya extract was treated as the therapeutic agent against AS & H2O2 in lane (7, 8) & (9, 10) (Fig. 4a).
Results of In vitro study of Catalase and SOD (Cu-Zn SOD or SOD1) activity by sodium arsenite with free phosphorus and cysteine. The present liver-slice experiment suggests that NaAsO2 (As3+) alone or in combination with H2O2 inactivates the SOD1 activity time-dependently Fig. 4d, lane 2, 3. Figure 4d, lane 4, 5, 6, 7, shows that free phosphorus and cysteine are found to be definitely SOD1 protecting agent. Catalase assay in the gel shows that the catalase activity is decreased by the As+ H2O2 & As in the lane (2, 3) respectively than the control (lane 1). The activity of the catalase was regained when the free phosphorus and cysteine were treated as the therapeutic agent against AS & H2O2 in lane (lane 4, 5, 6, 7) (Fig. 4e).
|
|
|
|
|
|
|
|
|
|
|
DISCUSSION |
|---|
In the present investigation, sodium arsenite (NaAsO2) treatment at a dose of 0.6 ppm/ 100 g b.w. for 28 days via drinking water to female Wister rats, a significant degeneration of the intestinal tissue is observed and that protection is noticed in the BBE supplemented group. The report suggests that arsenic, especially in its inorganic form, causes tissue death mainly by necrosis, apoptosis, cancers, and other abnormalities (29). Intestinal epithelial cells are the primary target to receive the ingested arsenic which is largely metabolically unmodified. In this connection, the present study is important. Ingested arsenic remains in inorganic form and the toxicity/DNA-breaking manifestations are mainly attributed by arsenite and arsenate in intestinal cells.
Arsenic exposure could induce the production of ROS which played critical roles in arsenic-induced toxicity (30-31). Lipid peroxidation generates a variety of relatively stable decomposition end-products, such as MDA, which can be measured as indirect indicators of oxidative stress (32). In this study, we found that the levels of ROS products such as MDA in intestinal epithelial tissues increased after arsenite exposure. Treatment of BBE significantly inhibited NaAsO2 induced ROS and MDA production. Arsenic-induced damage in the antioxidant system involves several mechanisms such as altered Superoxide Dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX) expression (33). In this study, our results showed that the inhibition of SOD and CAT levels by NaAsO2 was abolished by the treatment of BBE. These results indicated that BBE exhibited antioxidant effects in NaAsO2 induced intestine injury. Despite these extensive defense systems, biomolecule damage may still occur and persist within the cells and decrease the tissue integrity. The significant increase in the activities of SOD and CAT suggests a greater level of endogenous antioxidant associated with the BBE treatment resulting in an enhanced free radical scavenging activity (34). Small aquatic animals Bellamya extract (BBE) are the sources for a wide variety of compounds like ascorbic acid, thiol-containing amino acid, phosphorous etc (35). These compounds may be responsible for increasing antioxidant status.
Present results suggest that the two main causes of NPSH depletion were oxidative reaction with overproduced ROS and the arsenic methylation process. However, because GSH is almost always present in all cells at high concentrations (generally millimolar) (36), the two mechanisms alone do not seem to explain the striking depletion of NPSH in the tissue.
Arsenic exposure also exhibits oxidative stress through a significant reduction of GSH in liver, cultured lung epithelial cells and brain tissues (37). The NPSH, an antioxidant itself and a precursor of GSH are significantly declined by arsenic but that is restored by the flesh extract of Bellamya. The present protection by BBE has been demonstrated at the level of its antioxidative function in the restoration of several enzymes. That finally results in a protection in the macromolecular structures like DNA and proteins in the rat as noticed in the comet assay result. Arsenic-mediated induction of proinflammatory cytokines (IL-6 and TNF-α) is evident (38), which is noticed in the present study. Here, the BBE notably restored the TNF-α level. The elevation of metabolic inflammatory substance, C-reactive protein (CRP) has been noticed in an earlier study (39). Arsenic-induced alterations in nitric oxide (NO) in the blood and tissues have been reported and also evident in the present study (40). Alterations of nitric oxide (NO) in the periphery and systemic circulation have been detected after arsenic exposure. The NOS activity and NO metabolites content were also found to be lower. The decrease in NO level and nitric oxide synthase (NOS) activity may result in an inadequate NO signaling which has been correlated to abnormal cellular metabolism (40). Moreover, the generation of several free radicals (ROS) and nitrogen species during arsenic exposure may result in tissue degeneration, necrosis, and carcinogenesis (40-41). Anti-cancer activity of some natural products from marine organisms has been demonstrated. Commercial forms of the drugs prepared from these products are used in different physiological ailments (antitumor, analgesia, antiinflammatory, immunomodulation, allergy, and anti-viral) (42-43).
Arsenic exposure resulted in an elevation in total white blood cell (WBC) count with neutrophil and reduction in lymphocyte number (Table 1). Exposure of rats in this study to sodium arsenite produced leukocytosis. Increased number of WBCs is generally known to be a normal reaction to foreign substances. Leukocytosis, mainly due to leukocyte mobilization into the circulation can be a result of stimulation of the immune system against infectious agents or chemical (44). On the other way, it has been restored by BBE supplemented group (Table 2). From the results presented in, there is a significant increase in the activities of AST, ALP, ALT in the serum of arsenic-treated rats when compared to their control (Fig. 1f, 1g, 1h). This might have resulted from sodium arsenite-induced oxidative stress-related damages to hepatocytes membrane and leakage of hepatic transaminases into extracellular spaces (45). However, there was a decrease (P>0.05) in serum activities of AST, ALP and ALT in the NaAsO2+ BBE pretreated group compared to that of NaAsO2 only treated group. This suggests that hepatocytes are protected from arsenic by treatment of BBE. This can be attributed by the antioxidant present in BBE. In this experiment, sodium arsenite treatment in rat interfered with kidney functions as seen by elevation of these values. Urea, a waste product of protein catabolism, can rise when the kidney is defective. In renal disease, the serum urea accumulates because the rate of serum urea production exceeds the rate of its clearance (46). Increased urea and creatinine levels observed may be an indication of nephrotoxicity by sodium arsenite. This is in consonance with the findings of that arsenite toxicity induces several metabolic disorders including urea and creatinine elevation following proximal tubule damage and glomerular injury (47-48). Pretreatment with BBE had a reversal effects on these parameters. This study, therefore, showed the nephroprotective effects of the extract of bellamya in arsenite-induced toxicity.
In the present study, the cytosolic superoxidase dismutase activity was tested in gel zymogram study where it was noticed that several NaAsO2 treated group moderately decrease its activity in the liver cell. It has been extended higher in long duration incubation group but this impairment of SOD activity was successfully prevented by the BBE treated group. This activity increased then the control. Increased SOD activity is generally produced higher amount of H2O2 which is an inducing factor for catalase activity. In the present study similar pattern of response like SOD activity was noticed in catalase gel zymogram. The moderate decrease of catalase activity by As was efficiently restored and even increased in BBE treated group. This suggests that the combination protection of SOD & catalase in addition to thiol increase was more efficient for hepatic DNA protection.
From all these experimented results we can conclude that BBE has significant roles for the protection of cellular macromolecules like DNA and protein. And this protection was favorable to prevent against As-induced oxidative stress.
|
|
CONCLUSIONS |
|---|
Therapeutic efficacy study and reports on any mollusck and especially Bellamya sp. against metal toxicity are scanty. In this regard, the present work is of great value. Vast areas in South-East Asian Peninsula and other parts of the world are contaminated with arsenicals. A cheap, nontoxic and non-invasive natural therapeutic constituents or formulation is a demand of present time. The long term use of some natural substance which is enriched with nutritive value may offer least toxicity and side-effects. Our present data and our earlier published results strongly suggest that the aqueous extract of Bellamya bengalensis is potential against arsenic induced cellular toxicity. A large community of the globe consumes this benthic fauna as those are highly pertinacious. Our present outcome may be utilized for the development of some protective/therapeutic component against arsenic toxicity from this aquatic organism. Further studies are necessary for more conclusive comments.
|
|
CONFLICTING INTERESTS |
|---|
The authors declare that no conflicting interests exist.
|
|
ACKNOWLEDGEMENT |
|---|
University Grants Commission (UGC, MANF New Delhi) fellowship and contingent grant (awardees Sk. Sajed Ali).
|
|
REFERENCES |
|---|