| REVIEW ARTICLE |
|
|
Shantou Univ, Brain Funct & Dis Lab, Med Coll, Shantou, Guangdong, China
Corresponding Author: Shi-wan-er Chen, Shantou Univ, Brain Funct & Dis Lab, Med Coll, Shantou, Guangdong, China. E-mail: 22swechen@stu.edu.cn.
Running title: UNDERSTANDING THE MUSCARINIC ACETYLCHOLINE M3 RECEPTOR
|
|
ABSTRACT
|
|---|
Muscarinic acetylcholine M receptors (mAChRs) belong to the G protein-coupled receptor (GPCR) superfamily and play a crucial role in regulating and controlling the release of acetylcholine (ACh). This review aims to focus on M3 receptor (M3R) subtype distribution, function, efficacy, and clinical applications. Specifically, we will delve into the M3R, encoded by the CHMR3 gene, which is primarily found in the cerebral cortex, airway, digestive tract, and glands. Under normal healthy conditions, M3Rs participate in various significant physiological reactions. They are involved in regulating the release of second messengers and controlling the opening of ion channels. Such functions contribute to maintaining essential processes within the body. In pathological conditions, M3Rs become potential therapeutic targets. Pharmacological manipulation using M3R agonists or antagonists can be utilized in treating a range of diseases, including schizophrenia, Alzheimer's disease (AD), type Ⅱ diabetes (T2D), congestive heart failure (CHF), chronic obstructive pulmonary disease (COPD), acute pancreatitis (AR), and overactive bladder (OAB). These receptors hold promise as proven or potential targets for effective disease management. Therefore, understanding the pharmacological properties and molecular mechanisms of M3R is vital for developing multi-target drugs (MTAs) to treat diseases with complex pathogenesis. This approach can help avoid the decline in drug efficacy caused by drug resistance. Exploiting the unique three-dimensional structure of M3Rs and their selective preference for different ligands, selective or non-selective pharmaceutical reagents for targeted therapy can be designed. Understanding these characteristics can facilitate the design of drugs that interact optimally with the receptor, potentially enhancing therapeutic outcomes.
KEY WORDS: Muscarinic M3 receptor; G protein-coupled receptor; Physiological function; Pharmacological Effect; Therapeutic target|
|
INTRODUCTION TO MUSCARINE ACETYLCHOLINE RECEPTORS |
|---|
Introduction to acetylcholine (ACh) and Its Receptors
Muscarinic receptors serve as crucial targets for chemical signal transmission and play essential roles in various biochemical and biophysical events within the nervous system. They facilitate intercellular information exchange in the human body through signal transduction. Among the mammalian species, the G protein-coupled receptor (GPCR) superfamily is the largest group of receptors and is involved in nearly all physiological processes. Remarkably, about 30% of marketed drugs target GPCRs, making them significant in pharmacological research (1).
GPCRs possess structural characteristics, consisting of a single polypeptide chain with seven transmembrane segments (TM1-TM7). Each of these segments forms an α-helical structure and contains highly conserved amino acid residues. The center of the transmembrane helical segment, which exhibits significant homology (92%) (2) in amino acid identity, serves as the ligand binding pocket where GPCRs interact with their ligands. However, due to their high structural homology, targeting GPCRs through the orthosteric binding site (3) has proven challenging in obtaining subtype-selective agonists or positive allosteric modulators.
ACh is a neurotransmitter with multiple functions, playing vital roles in nerve excitability, skeletal muscle contraction, and essential central processes such as sleep, cognition, motor control, and sensory processing (4). Neurons that contain ACh are known as cholinergic neurons. The receptors that regulate cholinergic nerves fall into two main categories: muscarinic acetylcholine receptors and nicotinic acetylcholine receptors (nAChRs). Muscarinic acetylcholine receptors are found on effector cells innervated by parasympathetic postganglionic fibers, while nicotinic acetylcholine receptors are located on the postsynaptic membrane of both sympathetic and parasympathetic ganglion neurons, as well as the endplate membrane at the neuromuscular junction.
Muscarinic receptors belong to the rhodopsin family of GPCRs and are encoded by five genes (CHRM1-CHRM5). These genes are intron less within the coding regions and exhibit apparent similarities across mammals (5, 6). However, sequence diversity between species may lead to notable differences in ligand affinity (7). Upon activation, muscarinic receptors trigger canonical second messenger cascades, involving small non-protein molecules produced within the center neurons (8). In summary, ACh and its receptors, particularly the muscarinic receptors, play pivotal roles in various physiological processes. Understanding their structural characteristics and genetic variations is crucial for the development of targeted pharmacological interventions and the design of drugs that can selectively modulate these receptors for therapeutic purposes.
Introduction to muscarinic receptor subtypes and structure
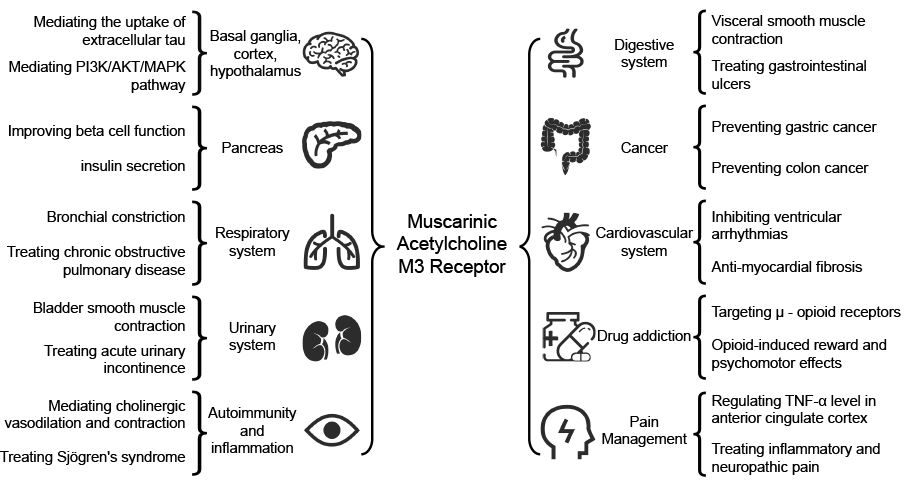
The selective coupling between receptors and ligands leads to biochemical reactions at specific sites, categorized as reaction sites and binding sites. When these sites are not in the same location, an excitatory effect is triggered, but if they overlap, an antagonistic effect occurs. The expression and function of different muscarinic receptor subtypes vary among tissues. Stimulating or inhibiting these subtypes can result in distinct therapeutic effects on disease remission (see Table 1). A concise summary of the characteristics, functions, and targeting of M3 receptors in health and disease has been provided (see Figure 1).
|
|
Muscarinic receptor subtype 1 (M1R). The levels of M1Rs are highest in the brain's cortical regions, making them the dominant approach for increasing neuronal excitability in the prefrontal cortex (20). In the Hepatic injury model, M1Rs can reduce the degree of liver fibrosis in mice, an effect that can be intensified by the activation of M3Rs.
Muscarinic receptor subtype 2 (M2R). In the brainstem and thalamus, M2 mAChRs are located at cholinergic synaptic terminals, controlling the release of ACh. In internal organs, smooth muscle contraction is modulated through a combined effect of M2Rs and M3Rs. M2Rs play a synergistic role and indirectly induce smooth muscle contraction by preventing cAMP-dependent relaxation in the presence of muscarinic agonists. On the contrary, M3Rs are chiefly responsible for cholinergic constriction of peripheral airways (43). There are also reports suggesting that the opening of non-selective cationic channels requires co-activation of both receptor types (44).
Muscarinic receptor subtype 3 (M3R). Although M3Rs are abundant in the central nervous system, they have no significant pharmacological effect due to weak expression. Instead, they are primarily found in the peripheral bladder, gastrointestinal tract, and glands. M3Rs, along with M4Rs and M5Rs, are present throughout the digestive tract, where they regulate the secretion of hydrochloric acid by parietal cells.
Muscarinic receptor subtype 4 (M4R). Similar to M1Rs, M4Rs can be targeted for mental disorders in brain neurons. However, the distinction lies in their enrichment in the caudate nucleus of the corpus striatum, where they primarily play a central excitatory role by activating dopaminergic neurons. M4Rs not only regulate basic cell movement but also prevent the autonomic inhibition of ACh in the heart and bladder.
Muscarinic receptor subtype 4 (M5R). M5 receptors have the lowest level among the five receptors, and their distribution is mainly limited to the hippocampus and VTA with discrete localization in the substantia nigra. Notably, blocking M5Rs has shown promise as a suitable approach in drug dependence and addiction, as seen in hypothalamic stimulation-induced reward effects in M5-knockout (KO) mice on morphine.
Odd-MR and Even-MR. The five types of muscarinic receptors can be categorized into two groups: MRodd includes M1Rs, M3Rs, and M5Rs, while MReven contains M2Rs and M4Rs. These classifications are based on the G protein types they preferentially couple with. The MRodd receptors are generally excitatory subtypes, primarily coupling via the α subunits of Gq/11 proteins (45), leading to increased nervous system excitability through activation of PLC and increased intracellular calcium concentration. On the other hand, MReven receptors preferentially couple with the Gi/o signaling pathway and are inhibitory subtypes. They mediate various physiological functions by reducing cellular cAMP levels (13) and inhibiting AC activity while prolonging the opening of potassium ions (12). They also control the opening of non-selective cation channels and transient receptor potential channels. Pertussis toxin-catalyzed adenosine diphosphate ribosylation can block the activation of MReven, but it is insensitive to MRodd.
Experimental methods for identifying MR functions. Previously, researchers compared pKb values between reagents and receptors to determine which receptor subtypes mediated a given functional response. More recently, specific KO mouse models have been created using gene editing technology, providing a more accurate way to quantify and judge the expression of each MRs by comparing normal wild type (WT) mice with gene-KO mutant mice (46-48).
|
|
INTRODUCTION TO M3 MUSCARINE ACETYLCHOLINE RECEPTORS |
|---|
M3Rs’ physiological roles in health conditions
M3Rs are widely expressed in different tissues of healthy experimental animals and humans (49), particularly in the urinary and gastrointestinal tract. In the PNS, M3 receptor activation regulates blood pressure (6), mediates smooth muscle contraction (50), and controls glandular secretion (51). In the CNS, M3 receptors are mainly concentrated in the brain, enriched in the dorsal vagal complex of the brainstem, arcuate nucleus, and ventromedial hypothalamic nucleus. However, their expression level is lower compared to M1 and M2 receptors. M3Rs play a vital role in neurotransmitter release (52), motor control, temperature homeostasis, food intake and weight regulation (53), cardiovascular regulation (54), and memory, indicating their essential involvement in regulating a broad range of normal physiological processes.
The activation of second messengers is linked to the mobilization of the phosphoinositide system in hormone information transmission. When ACh, released from the end of parasympathetic nerves, binds to the specific M3 receptor on the cell membrane as the first messenger, it couples with Gq/11, one of the G protein subtypes. This activation then triggers phosphatidylinositol-specific PLC (PI-PLC) on the plasma membrane, leading to the release of two intracellular second messengers, DAG and inositol 1,4,5-triphosphate (InsP3) (55, 56). Muscarinic agonists can also attenuate potassium currents by decreasing the amount of phosphatidylinositol-4,5-bisphosphate (PIP2) (57). DAG, a substrate for the synthesis of phosphatidic acid, has the effect of inhibiting weight gain and lowering blood lipid levels. InsP3 tightly binds to the endoplasmic reticulum (ER) receptor and changes its conformation, leading to the opening of calcium channels and participating in the regulation of extracellular calcium influx. This results in increased mobilization of Ca2+ from intracellular storage and reduced intracellular Ca2+ overload (58-60).
Calcium ions at physiological concentrations combine with DAG to activate Ca2+-dependent PKC. PKC, belonging to a subgroup of the highly conserved serine/threonine kinase family, is involved in phosphorylation of target proteins and regulates various signal pathways. Activated PKC can control a range of physiological processes, including redox signaling, oxidative stress disorder, gene expression, tissue damage repair, vesicle transport, cell proliferation, differentiation, migration, and apoptosis (61-63).
Apart from their effects on smooth muscle contraction and glandular secretion through the InsP3-Ca and DAG-PKC signal transduction pathways, M3 receptors also activate downstream key protein kinases by regulating other pathways such as the Wnt pathway (64) and the epidermal growth factor receptor (EGFR) pathway (35, 65).
The interplay between M3 receptors and these pathways significantly impacts cell life progression and ultimately reduces the metastatic rate and invasiveness of cancer cells. For instance, the activation of M3 receptors in endothelial cells expressing MMP-1 and MMP-7 catalyzes the release of heparin-binding EGF-like growth factor (HB-EGF), leading to the activation of the EGFR pathway and promoting cancer invasion through downstream cancer gene transcription via the ERK 1/2-phosphatidylinositol-3-kinase (PI3K) pathway (66).
In a normal physiologic state, vascular smooth muscle endothelial cells promote the production of endothelial nitric oxide synthase (NOS) upon receiving signals from M3 receptor activation. This increase in nitric oxide (NO) concentration inhibits the inward flow of calcium ions and the outward release of calcium ions from the intracellular calcium pool, leading to the relaxation of vascular smooth muscle fibers, beneficial in reducing vascular function damage and protecting endothelial cells (67).
In conclusion, in healthy conditions, M3 receptors, upon binding to specific ligands, initiate a series of physiological activities mainly through InsP3-Ca and DAG-PKC signaling pathways. They are actively involved in tumorigenesis and tumor inhibition, myocardial protection, taste regulation, smooth muscle contraction, blood glucose regulation, vesicular exocytosis, and other biochemical reactions.
M3Rs' Pharmacological Role in Pathological Conditions
Drugs targeting M3R have drawn considerable attention from medical researchers for a long time. Given the widespread expression of M3R in both central and peripheral regions and its significant pharmacological effects in regulating the release of ACh, understanding the potential molecular mechanisms of M3R is challenging but essential in identifying new therapeutic targets for clinical treatment of various pathological conditions. These conditions include neurological diseases and mental disorders (e.g., mood disorders, psychosis, anxiety, and dementia), endocrine system diseases (e.g., Type Ⅱ diabetes mellitus, SS), circulatory system diseases (e.g., congestive heart failure, arrhythmia), respiratory system diseases (e.g., chronic obstructive pulmonary disease, asthma), digestive system diseases (e.g., stress-induced acute response, colorectal cancer), and urinary system diseases (e.g., overactive bladder).
M3Rs in Neurological and Psychiatric Disorders. Given the widespread projections of cholinergic neurons in the central nervous system, it is well-established that muscarinic receptors, mediating cholinergic transmission, play a significant role in the pathophysiology of neurological and psychiatric disorders. The transmission of cholinergic nerve signals is significantly impacted by the muscarinic receptor and nicotine receptor families, both of which play essential roles in the pathophysiological processes of mental and neurological disorders. The current study extensively utilizes animal and cell experiments to investigate the targeting of muscarinic receptors for the treatment of these disorders. These receptors hold substantial promise as potential targets for the development of therapeutic drugs in the future (68). Numerous studies have shown that the muscarinic M3 receptor subtype, mainly expressed in the hippocampus and cerebral cortex, plays a crucial role in the cognitive, memory, and learning functions of individuals with psychiatric disorders. As such, it is considered a therapeutic target for various psychiatric disorders, including schizophrenia, bipolar disorder, manic-depressive psychosis, as well as for neurological diseases, including Alzheimer's, Huntington's, and Parkinson's diseases.
Alzheimer's disease (AD) (69) is a chronic neurodegenerative disease primarily affecting elderly individuals over the age of 60. During the disease's progression, neurons related to learning and memory in the brain are gradually damaged or lost, leading to clinical symptoms such as short-term memory loss, progressive cognitive decline, and non-cognitive mental symptoms, ultimately impairing social functioning. Recent reports have indicated that the level of M3 receptors in the submandibular gland of AD patients is decreased compared to non-dementia healthy controls, while the level of ACh is found to be increased. This suggests that the M3 signal pathway mediated by ACh in the salivary gland of AD patients is impaired, leading to salivary gland dysfunction and reduced secretion of salivary lactoferrin (70). Given the unclear pathogenesis and the lack of a radical therapeutic schedule, M3 receptor may represent a suitable alternative therapy for improving AD-related memory loss (68, 71).
Characteristic components of nerve fibers in brain cells of patients with neurodegenerative diseases are microtubule-associated protein tau (MAPT) (72) tangles. The content of phosphorylated tau (p-Tau) protein, which experiences function loss due to abnormal post-translational modification, is significantly increased in the brain of patients with AD (73). This suggests that tau protein levels are closely related to cognitive dysfunction (74). M3 receptors can mediate the uptake of extracellular tau and then combine with tau to increase the intracellular calcium level of neurons (75).
Cholinesterase inhibitors form the cornerstone of Alzheimer's disease treatment as they indirectly make M3 receptors a potential therapeutic target for AD (76). For example, the pharmacological mechanism of Huperzine A is to indirectly increase the level of endogenous ACh by inhibiting cholinesterase (77). Subsequently, the activated M3 receptor acts on the protein kinase B (PKB/AKT)/mitogen-activated protein kinase (MAPK) pathway, exerting a neuroprotective effect on the treatment of benign memory disorders. Traditional Chinese medicine research has indicated that numerous Chinese herbal extracts and decoctions, such as Xixin Decoction (78), geraniol (79, 80), Tinospora sinensis (81), resveratrol (82), and Andrographis paniculata (83), can positively impact the treatment of AD by mediating the downstream PI3K/AKT/MAPK signal pathway.
Schizophrenia is a prevalent and debilitating mental illness, characterized by chronic persistence and a profound disconnect between a patient's mental state and their environment, leading to a substantial deterioration in their quality of life. It comprises a wide range of symptoms and may lead to complications such as tuberculosis, heart disease, and liver disease. According to research, specific subtypes of receptors—namely, M1, M2, and M4—are associated with schizophrenia and influence the effectiveness of antipsychotic drugs. In particular, M1 and M4 receptors show promise as potential targets for treating schizophrenia (84). Notably, drugs that target these receptors not only exhibit antipsychotic effects but also have the potential to ameliorate cognitive and motor dysfunction.
However, as indicated in the table above, M3 receptors are predominantly found in the peripheral nervous system, which may limit their impact on the central nervous system—the main factor hindering their involvement in schizophrenia treatment. Neuroimaging studies focused on protein expression reveal lower levels of M1/M4 receptors in the cortex and subcortical regions of individuals with schizophrenia. Moreover, in the hippocampus, there is a reduced binding density of ligands interacting with M4 receptors. Alterations in M2 and M3 receptors are primarily confined to subcortical regions. The distribution pattern in the body suggests that the M3 receptor may not play a pivotal role in schizophrenia treatment.
M3Rs in Type II Diabetes Mellitus. When clinically treating psychiatric disorders with antipsychotic drugs like olanzapine, clozapine, and aripiprazole, there may be serious metabolic side effects such as weight gain and glycolipid metabolic disturbances due to M3 receptor signaling blockade. This suggests that M3 receptors may play a fundamental role in regulating glucose and lipid metabolism balance, insulin secretion, and energy balance in the brain. Research on human pancreatic islets indicates that ACh, synthesized, stored, and released by pancreatic α cells, can transmit paracrine signals to adjacent β cells, activating M3 receptors on β cells and promoting glucose-stimulated insulin secretion (GSIS). As such, the M3 receptor on β cells is considered a potential target for drugs that promote insulin release.
Type Ⅱ diabetes mellitus (T2DM) is a common chronic metabolic disease characterized by the inability of pancreatic β cells to release sufficient insulin to cope with peripheral insulin resistance (85). Under pathological conditions, patients with diabetes often present with dysglycemia, dyslipidemia, obesity, polyphagia, polydipsia, polyuria, and weight loss. Numerous studies have demonstrated that promoting insulin release and regulating leptin are the primary points of the pharmacological mechanisms of pancreatic β cell-expressed acetylcholine muscarinic M3 receptor in improving obesity symptoms and treating diabetes. Mutant mice models suggest that M3 receptors significantly contribute to the normalization of pancreatic β cell function. Mice with M3 gene knockout develop a phenotype characterized by impaired glucose tolerance and significantly reduced insulin release. These animal experiments provide convincing evidence that M3 receptors are closely linked to glucose metabolism capacity and glucose homeostasis (86, 87).
ACh, released by pancreatic α cells, stimulates M3 receptors expressed by pancreatic β cells, leading to an increase in glucose sensitivity of β cells (88). Under pathological conditions, M3 receptors work to restore impaired glucose control and regulate blood glucose normalization. Specifically, M3 receptors can promote glucose-stimulated insulin secretion in diabetic animal models, and conversely, inhibit insulin overproduction in atropine-induced animal models (89).
In 2020, over 50% of Chinese adults were overweight or obese, making China one of the world's most obese nations. Researchers are increasingly concluding that insulin resistance, glucose lipid metabolism problems, and obesity are interconnected. The M3 receptor is involved in numerous processes related to the regulation of lipid and glucose levels, which can help alleviate some symptoms caused by lipid or glucose imbalances. Obesity symptoms in diabetes patients may be associated with asthma occurrence. The M3 receptor on airway smooth muscle cell membranes, when stimulated by leptin, a product of obese gene-mediated adipocyte secretion (90), can inhibit parasympathetic signaling, relax bronchi, regulate lung function, and contribute to the treatment of obesity-associated asthma. Leptin hormone signaling mediates processes such as metabolism, appetite suppression, and sugar and lipid reduction (91). Pancreatic β cell-specific M3 receptors can increase the expression of leptin genes and enhance the synthesis of leptin receptor proteins in the hypothalamus. Obesity is related to impaired salivary secretion; however, activation of M3 receptors can promote hormone secretion from salivary and submandibular tissues, thus improving the function of secretory structures or glands. M3 receptors can affect leptin gene expression and increase hypothalamic leptin receptor protein synthesis.
M3Rs in Respiratory Diseases. Chronic obstructive pulmonary disease (COPD) is a chronic airway inflammatory disease characterized by severe destruction of the normal lung morphology. It includes chronic bronchitis and emphysema and is distinct from bronchial asthma due to its incompletely reversible airflow limitation. Bronchodilators like salbutamol and ipratropium bromide, which target M3R and β2-adrenergic receptors (β2Rs), are commonly used for COPD stable treatment.
M3R and β2Rs are essential regulators promoting intracellular calcium release and improving airway cell function. First-line drugs for treating COPD, asthma, and obstructive pulmonary disease often target and bind to these receptors, playing a crucial role in mediating the pathological contraction observed in asthma and COPD. Short-acting β2 receptor agonists (SABA) (92) and short-acting muscarinic antagonists (SAMA) are limited to symptomatic treatment of acute bronchial asthma exacerbations in clinical studies of respiratory diseases. On the other hand, long-acting muscarinic antagonists (LAMA) (93) are of great value in both monotherapy and combination therapy for maintenance treatment and exhibit myriad usefulness in the treatment of lung diseases. LAMA represents a significant milestone in the treatment of bronchial asthma due to its ability to efficiently and selectively block the combination of ACh and muscarine receptors, thereby inhibiting parasympathetic nerve impulses. For example, LAMA can be used as a bronchodilator in the treatment of COPD, asthma, and other respiratory diseases, as demonstrated by in vitro and in vivo studies on ACh receptor-KO mice. The effect of LAMA extends to anti-inflammatory properties, both alone and in synergistic combination with long-acting β2R agonists, as evident in the phenotype of COPD model animals. The synergy between M3 muscarine receptor antagonists and β2R agonists is achieved through cross-regulation or crosstalk between M3R and β2R, ultimately determining the contraction state of airway smooth muscle (ASM), airway diameter, and airflow resistance.
Among the main muscarinic receptor subtypes distributed in the airways, urinary tract, and gastrointestinal tract, the M1, M2, and M3 receptors stand out. Particularly, the M3 receptor is considered the main mediator of ASM contraction or tension under physiological conditions (94). It plays a significant role in relaxing airway smooth muscle, improving lung function, reducing mucus secretion, and attenuating airway hyperresponsiveness. Conversely, M2 receptors antagonize G protein-induced bronchodilation and drive airway smooth muscle contraction through an indirect feedback mechanism (95, 96). Blockade of neuronal M2 receptors in the airways increases ACh release, decreases cAMP levels, and reverses vagal nerve-induced bronchial reflex constriction. Additionally, M3 receptor antagonists activate the PLC-IP3-Ca2+ signaling pathway through coupling with Gq (97). This activation not only directly facilitates smooth muscle contraction and airway tone but also suppresses symptoms of bronchial hyperconstriction, mucus hypersecretion, and airflow limitation in asthmatic patients. These effects have been confirmed through diverse clinical trials as effective and efficient therapeutic interventions for COPD and asthma. A recent study explored the mechanism of inhibiting M3 receptor to improve airway inflammation by using the M3 receptor antagonist thiamethoxam in asthma model mice. The experiment found that thiamethoxam plays a preventive role in allergic airway inflammation by inhibiting the development and differentiation of M2 macrophages (98). However, blocking M2 receptors in the airway may induce an increase in ACh release and promote bronchoconstriction, thereby diluting the effect of M3 receptor antagonists on smooth muscle relaxation (99). To address this, drug development has focused on enhancing the M3 receptor selectivity of muscarinic antagonists and reducing their residence time at M2 receptors, aiming to decrease the incidence of cardiovascular system side effects. Recent studies have shown that representative long-acting antimuscarinic agents such as revefenacin (100), imidafenacin, glycopyrrolate (101), tiotropium (102), and aclidinium (103), which have high antimuscarinic activity and M3 subtype selectivity, exhibit good long-lasting efficacy, tolerability, and safety profiles, effectively improving the quality of life for patients. The choice of suitable bronchodilators depends on specific conditions in clinical practice.
M3Rs in Urinary System Diseases. Overactive bladder (OAB) is a urinary dysfunction syndrome characterized by symptoms such as urgency and urge incontinence, first identified in 1971. Urodynamic studies describe it as an "unstable bladder," with non-inhibitory contractions of the detrusor muscle and frequent urination intentions.
Around 50% - 60% of M3 receptor complexes are associated with the plasma membrane of the human bladder detrusor muscle (104). This may explain that the genesis of OAB involves excessive contraction and decreased relaxation of the bladder detrusor smooth muscle due to M receptor overactivation. The research on bladder diseases has predominantly focused on M2 receptors in the transverse muscle layer and M3 receptors in the longitudinal muscle layer of the urethra (105). The latter is considerably more responsible for the contractile reaction after being stimulated. M2 receptors indirectly induce relaxation by blocking cAMP stimulation and play a secondary synergistic role in improving bladder dysfunction. However, they cannot participate in the M3 receptor-mediated contraction in vitro. Therefore, the mainstream therapy for OAB is anticholinergic therapy, which involves inhibiting M3 receptors located in the urinary tract epithelium that mediate bladder contraction (106). Under pathological conditions, M3 receptor antagonists directly increase the excitability and contractility of bladder smooth muscle by inhibiting the activity of the big conductance calcium-activated potassium (BK) channel (107) and promoting the opening of L-type voltage-dependent calcium (VDC) channels through excitation-contraction coupling.
Researchers have attached utmost significance to developing new formulations with high bladder selectivity to improve the efficacy and reduce adverse drug reactions (ADRs) of OAB therapeutics. Solifenacin succinate (108, 109), a new breed of M3 receptor antagonist, primarily inhibits bladder detrusor asymmetric or irregular contractions. It is considered a first-line drug for adult OAB due to its high accuracy in targeting the bladder and having fewer potential side effects on the brain and heart compared to nonselective antagonists, although it may still generate ADRs such as constipation, dry mouth, urinary tract infection, and severe anaphylaxis. Another novel anticholinergic, dalfampridine, which has a highly precise positioning for the M3 receptor, effectively increases bladder reservoir volume and reduces the frequency of urine. Similarly, the therapeutic approach of altering M3 receptor activity may be effective in treating neurogenic detrusor overactivity (NDO) (110). Drugs like Mirabegron (111) sustained-release tablets and oral suspension based on M3 receptor antagonist mechanisms are approved for treating pediatric patients with NDO. Other drugs like oxybutynin in transdermal administration, suitable for female patients with OAB, and midanacin with oral administration not only convey high affinity to M3 receptors but also exert long-lasting efficacy and safety with fewer ADRs, such as dry mouth, stomachache, and constipation. Additionally, oral or intravenous tolterodine (112), a highly bladder-selective M3 receptor antagonist, reduces bladder wall thickness and increases bladder capacity. Fesoterodine (113, 114), a first-line therapeutic agent for OAB, exhibits high affinity for M2 and M3 receptors and does not enter the blood-brain barrier, providing rapid efficacy and fewer side effects.
M3Rs in Digestive System Diseases. M3 receptors play a crucial role in mediating immunity in inflammatory pathological states, such as primary biliary cholangitis, primary sclerosing cholangitis, allergic airway inflammation, and severe acute pancreatitis (SAP) (115).
Pancreatitis is an inflammatory reaction caused by the activation of pancreatic enzymes in the pancreas, characterized by pancreatic swelling and inflammatory cell infiltration in mild cases, and tissue hemorrhage, necrosis, infection, and shock in severe and acute lesions. Experiments with CHRM3-KO mutant mice insensitive to ACh indicate that M3 receptor activation is involved in the pathogenesis of pancreatitis.
In vitro experiments have shown that high concentrations of M3 receptor agonists can increase trypsin activity, possibly leading to pancreatic acinar cell injury. Conversely, M3 receptor antagonists can attenuate pancreatic mucosa damage by curbing enzyme activity in secretory fluid, preventing self-digestion of pancreatic tissue, and reducing pathological damage. M3 receptors, widely distributed in the digestive tract, especially in the pancreatic islet and acinar cell membranes (116), participate in pepsinogen secretion, vasoconstriction, oxidative stress, cell proliferation, and inflammatory signal pathways (117). Under pathological conditions, M3 receptor expression is related to the release of many inflammatory factors. Clozapine-n-oxide (CNO) (118), a ligand for the M3 receptor (119), induces prevacuolar cell death and inflammation, and M3 receptor antagonists ameliorate CNO-activated pancreatitis symptoms in WT mice. M3 receptors have a significant regulatory effect on the pathological damage of L-arginine-induced pancreatitis, particularly in regulating necrosis rather than apoptosis of acinar cells, thus playing a protective role against injury (120). Their rich physiological functions make them attractive potential targets for nonalcoholic fatty liver disease, cholangitis, and liver fibrosis (121). Blocking M3 receptors can lessen the severity of pancreatitis (122).
Based on theoretical basis and experimental validation, there is sufficient evidence to support that regulating M3 gene expression in the pancreas of SAP animals may positively influence preventing necrotizing pancreatitis under pathological conditions. Inhibiting M3 gene expression may be an effective and promising therapeutic strategy to prevent pathological necrotizing pancreatitis.
M3Rs in Autoimmunity and Inflammation. Research has demonstrated that M3 receptors play a crucial role in immune regulation and inflammatory responses. Modulating the M3 receptor holds potential therapeutic benefits for autoimmune conditions like Sjogren's syndrome and various inflammatory disorders. The M3 receptor primarily mediates cholinergic vasodilation and contraction, and its dysfunction can lead to vasoconstriction and hypoxemia (123). Impaired circulation and oxygen supply can result in numerous symptoms associated with various inflammatory conditions, such as encephalomyelitis.
Numerous experiments have highlighted the involvement of the M3 receptor in the immune response to airway pathogens, where it transmits stimulation signals to enhance mucus and cilia clearance rates (124). This receptor also acts as an autoantigen in autoimmune salivary gland inflammation and SS-like autoimmune sialoadenitis (125). For instance, experiments involving uninfected and Nippostrongylus brasiliensis-infected wild-type and type 3 muscarinic receptor (M3R)-deficient (CHRM3-/-) mice have demonstrated the positive role of M3 receptors in promoting mucosal homeostasis, eliminating intestinal pathogens, and regulating immune cell function (126). Furthermore, experiments have elucidated the M3 receptor's role in modulating macrophage phenotype and function. Additionally, activating M3 receptor activity can facilitate macrophage development, reduce goblet cell mucus secretion, and thus contribute significantly to defense against Citrobacter Rodentiae (127).
Sjogren's syndrome (SS) lesions are related to the inflammatory response and immune factors associated with ACh signaling transmission. The pathophysiology of SS involves T cells recognizing antigens through T cell antigen receptors, thereby triggering the production of cytokines and chronic inflammation (128, 129). It is characterized by lymphocytic infiltration, ductal dilatation or stenosis in the salivary and lacrimal glands, with typical manifestations including xerostomia and ocular dryness or xerophthalmia. The expression of CHRM3 mRNA in patients with primary Sjogren's syndrome was significantly higher than that in the healthy control group, and M3R responsive IL-17 secreting cells were also significantly increased (130).
A plethora of experimental data and academic literature explain the modifying effects of M3 receptors on SS symptoms, and what takes effect in this process are M3 receptor agonists. A potential treatment for SS is the M3 receptor agonist pilocarpine (131), which works by directly stimulating the gland's M3 receptor, increasing the expression of the M3R gene and protein, and promoting the secretion of tears and saliva by the lacrimal and salivary glands, respectively, thereby improving lacrimal gland dysfunction.
White peony, namely Shaoyao, a traditional Chinese herbal remedy that helps treat Sjogren's syndrome, contains a primary component β-Sitosterol that may increase the expression of CHRM3 to cause salivation to be secreted (132). The Shaoyao Gancao Tang (SGD) made from white peony and licorice can also regulate the CAMP-PKA signaling pathway, increase M3R levels, and improve SS symptoms (133).
In brief, SS is an autoimmune disease, and one of the autoantigens associated with SS is the M3 receptor (134), which is found in the exocrine gland with disease-specific inflammation. Antigen-specific targeted therapy can be used to target M3 receptors since M3R-reactive B cells and Th1 cells may have a pathogenic function in SS.
M3Rs in Cancer Treatment. The M3 receptor is expressed in specific cancer cells, and its activation is associated with tumor growth and metastasis. Targeting the M3 receptor offers a novel approach to cancer treatment.
Extensive research has highlighted the interplay between the cholinergic system and the immune system. The activation of M3 receptors is linked to cell mutation, division, metastasis, and proliferation in various types of tumors, including brain tumors, pancreatic tumors, and ovarian tumors (135). Animal experiments have revealed that M3R activation promotes the progression of gastric cancer, pancreatic cancer, colon cancer, as well as liver injury and fibrosis.
Experimental data using the murine colon cancer cell line CT-26 in vitro and an orthotopic mouse model of colorectal cancer in vivo demonstrated that the M3 receptor blocker 4-DAMP significantly inhibited CT-26 cell proliferation and induced apoptosis in a dose-dependent manner (136). The use of selective agonists and antagonists for different MRs has distinct therapeutic implications. Through G(q) and downstream activated M receptor signals, these compounds can initiate phospholipid turnover, modify cellular calcium levels, activate protein kinases, and ultimately influence gene transcription and cellular function.
M3 receptors are widely distributed in the gastrointestinal tract and liver, with a significant increase in M3 mAChR levels observed in gastric tumor tissue. Carcinogenesis may be promoted by the overexpression of nerve growth factor (NGF) via cholinergic stimulation in gastric epithelial cells. Research indicates that knocking out the CHRM3 gene encoding the M3 receptor can inhibit the proliferation of muscarinic M3 receptor-dependent epithelial cells, effectively impeding the formation of intestinal tumors (137).
The progression of prostate cancer is linked to the M3 receptor-induced targeted FAK-YAP signaling axis, and blocking muscarinic signaling can counteract the M3R-induced castration-resistant growth of prostate cancer cells (138, 139).
A large amount of literature elaborates that activation of M3 receptors is associated with cell mutation, division, metastasis, and proliferation in many tumors, such as brain tumors, pancreatic tumors, and ovarian tumors (140). Colorectal cancer (CRC) is a common malignancy of the digestive system with high morbidity and mortality. Multiple validation approaches in vivo and in vitro reveal that only the M3 subtype, one of the five muscarinic subtypes, is expressed in a massive number of colon cancer cell lines. Based on the overexpression of M3 receptor mRNA in CRC tissues and the fact that administration of muscarinic receptor agonists increased both the number and volume of CRC cells, it can be hypothesized that M3 receptors are affiliated with the cellular program that promotes the malignant or benign growth, survival, and spread of CRC (141, 142). In vivo and in vitro studies on colon cancer model mice have shown that using genetic or pharmacological methods to block the expression and activation of M3 receptors can weaken the occurrence and progression of colon cancer, respectively (143).
The pharmacological mechanism of ammonium bromide against CRC is that selectively restraining expression or blocking the post-signaling of M3 receptors can reduce the release of matrix metalloproteinase (MMP), which corresponds considerably to the growth of tumors, especially MMP-1 and MMP-7, to cut down the invasiveness and reverse the metastatic spread of CRC cells (144-146). Apart from that, it can lower the expression of the anti-apoptotic Bcl-2 protein and increase the levels of pro-apoptotic caspase-3 and Bax proteins. Conclusively, ammonium bromide efficiently inhibits CRC cell proliferation, prevents them from escaping programmed death, and accelerates their death (147). Immunohistochemical analysis of M3 receptor selective antibodies also supports the above statement, suggesting that M3 receptor has potential clinical application in CRC treatment (148).
M3Rs in Cardiology. Congestive heart failure (CHF) is a syndrome characterized by circulatory congestion, dyspnea, and limb swelling as a result of cardiac hypofunction. Mechanisms for treating CHF clinically include increasing cardiac output, reducing cardiac load, and reversing cardiac hypertrophy.
M3 receptors are enriched in gap junction channels between myocardial smooth muscle and exert anti-heart failure effects by regulating many factors in the circulatory system (149). For example, microRNAs are important messengers that regulate cardiac ion channel function and expression, and gap junction channel proteins have a strong association with cardiac injury and cardio-protection. The latest research has found that activated M3 receptors in the heart can directly target collagen to resist myocardial fibrosis by mediating the miR-29b/BACE1 axis, thereby protecting the heart (150).
In the cardiovascular system, the activated M3 receptor reduces the expression of microRNA-1, which is associated with myocardial hypertrophy. On one side, this effect increases InsP3 release, alleviates oxidative stress induced by calcium overload, inhibits the lipid peroxidation (LPO) process, and hinders the occurrence of atherosclerosis. On the other side, it improves myocardial contractility, increases cardiac output, and improves hemodynamics. M3 receptors enhance the expression of Cx43 (151), a gap junction channel protein relevant to heart failure, thereby exerting therapeutic effects in protecting cardiomyocytes from ischemic injury and reperfusion injury (152, 153). Animal model experiments further validate that the protective regulatory mechanisms of M3 receptors on the myocardium include the reversal of cardiac hypertrophy and resistance to myocardial and vascular remodeling, suggesting that M3 receptors can be potential therapeutic targets for chronic cardiac insufficiency.
Arrhythmia is a serious cardiovascular disorder characterized by abnormal heart frequency and rhythm due to blood supply insufficiency and conduction blocks. Clinical approaches to treating arrhythmias focus on reducing myocardial automaticity, posterior depolarization, and eliminating reentry, as well as shortening the action potential duration (APD) and prolonging the effective refractory period (ERP).
Reduced expression of the human ether-à-go go related gene (hERG) (154) leads to impaired electrical-triggered cardiac repolarization and an increased risk of fatal arrhythmias, contributing significantly to sudden cardiac death. Activation of cardiac M3 receptors can accelerate repolarization of the transmembrane potential and significantly shorten the APD, achieved by downregulating miR-1 expression and upregulating hERG levels. M3 receptors also promote the opening of inward rectifier/inwardly rectifying potassium (Kir/IRK) channels and the closing of calcium channels by inhibiting miR-1, resulting in increased potassium current on the cardiomyocyte membrane and inhibition of calcium overload. Upregulating M3 receptor expression through the dephosphorylation of Cx43 improves cardiac electrical signaling and reduces the occurrence of barium chloride-induced arrhythmia. Experiments on mice have shown that the non-selective cholinergic drug pilocarpine, acting on M3 receptors, significantly slows heart rate (HR), delays the onset of ventricular arrhythmia triggered by acute myocardial ischemia, and prolongs animal survival (155).
Cholinergic signaling mediated by M3 receptors has been linked to hypertension, and M3 gene-knockout reduces blood pressure salt sensitivity in hypertensive mice (156), leading to a drop in blood pressure (157). Additionally, reducing the gene expression of M3 receptors distributed in angiotensin Ⅱ (Ang Ⅱ)-induced vascular smooth muscle (VSMC) inhibits cell proliferation and migration, reverses vascular allostery, and improves vascular function (158). Partial blockade of M3 receptors has been found to increase nitric oxide and decrease miR-1 in the pulmonary circulation, suggesting a molecular mechanism by which it suppresses hypertension (159). When combined with ligands, M3 receptors have beneficial effects in relieving pulmonary hypertension (PH) complicated by right heart failure and improving pulmonary circulatory dysfunction by increasing NO production in blood vessels, repressing miR-1, and dilating pulmonary vasculature. Blocking the expression of M3 receptors on the cardiomyocyte membrane can modulate abnormal electrically triggering activity through delayed rectifier K+-current, thus ameliorating the harm caused by increased blood pressure and cardiac dysfunction. An in vitro isolated rat aortic rings assay illustrates that 4-hydroxybenzaldehyde, a component of antihypertensive Chinese medicine, exerts vasorelaxant effects through the M3-dependent PLC-IP3 pathway, as well as potassium and calcium channels (160). Inhibiting M3 receptors can be a valuable approach to investigate the underlying mechanism of antihypertensive drugs.
Despite considerable research efforts on the mechanisms involving M3 receptors in cardiovascular illnesses, there are still many molecular mechanisms to be discovered, validated, and thoroughly explained in the future.
M3 Receptor-Related Drug Addiction. In the field of neuroscience, addiction is widely recognized as a chronic and recurrent brain disease, characterized by individuals who continuously seek out potentially addictive substances without self-control. Addiction is closely related to brain regions that regulate reward circuits. Scientific studies indicate that addiction results in a decrease in the capacity of reward circuits to respond to rewards and a heightened sensitivity of emotional circuits to stress. This leads to a deficiency in self-regulation ability, particularly in the prefrontal cortex. People with addiction often experience stronger negative emotions and intense drug cravings, which can make it difficult to quit even with a strong will to do so (161).
The imbalance of dietary behavior shares similarities with compulsive drug consumption in addiction, as both involve the stimulation of reward pathways. The release of melanin concentration hormone (MCH) neurons in response to ingested food glucose triggers the reward system, promoting the release of dopamine in the brain. Foods rich in sugar and fat can particularly trigger this process, leading to increased dopamine production and stronger pleasure (162).
The mesolimbic dopamine system plays a central role in drug reward, and endogenous opioid drugs increase dopamine release in the nucleus accumbens (NAc) by acting on different molecular targets. Endogenous opioid peptides and their receptors, including μ (MOR), δ (DOR), and κ (KOR) opioid receptors, contribute to reward and analgesia, anxiety relief, and recurrence, respectively. MOR in specific brain regions is associated with the rewarding effect of opioids (161, 162).
The M3 muscarinic receptor is involved in opioid-induced reward and psychomotor effects. By co-localizing with μ-opioid receptors, it exerts inhibitory effects on GABAergic neurons in the brain, contributing to reward effects. This makes the M3 receptor a potential target for the treatment of opiate addiction. Studies have shown that the M3 receptor agonist pilocarpine inhibits morphine-induced conditioned position preference (CPP) (163), while the M3 receptor antagonist 4-DAMP reverses this inhibitory effect. The complex role of the muscarinic system in addiction pathophysiology suggests that M3 receptors and other muscarinic receptors may be linked to an increased risk of alcohol and cannabis dependence (163-165).
M3 receptor in Pain Management. Pain is an unpleasant physical and emotional sensation triggered by actual or potential tissue damage, and it can be categorized based on factors such as duration and location. The currently available scientific literature provides a robust theoretical framework for managing chronic non-cancerous pain. Research into pain treatment methods is ongoing because different types of pain require diverse approaches.
The cingulate gyrus in the brain is a crucial component of the limbic system and the medial pain perception system, influencing emotions, cognitive processing, attention, and movement. The anterior cingulate cortex (ACC) (166, 167) is involved in emotional autonomic responses, particularly in neuropathic pain (168), while the posterior cingulate cortex (PCC) does not play a role in these functions. The interaction between tumor necrosis factor-α (TNF-α) and neurons in the ACC may regulate the cytokine microenvironment, contributing to neuropathic pain.
Gq-coupled human M3 muscarinic receptors (hM3Dq), a modified form of human M3 muscarinic receptors (hM3), are exclusively activated by designer drugs (DREADDs), specifically clozapine N-oxide (CNO), and coupled through the Gq signaling pathway (169). Extensive clinical and experimental studies suggest that M3 receptors are associated with pain management and regulation. M3 receptors play a role in neuropathic pain by influencing the TNF-α level in the ACC and the mechanical paw withdrawal threshold, indicating a potential mechanism for pain management via hM3D receptors (170).
In an experiment based on Cre recombinase expression in transgenic mice driven by the vesicular glutamate transporter-2 (VGluT2) promoter, CNO-activated excitatory hM3D receptors significantly increased neuronal discharge and synaptic glutamate release. These findings suggest that targeting glutamatergic dorsal horn neurons with inhibitory DREADDs may offer a novel approach to treating inflammatory and neuropathic pain. Another study using a primary dysmenorrhea rat model demonstrated that pentylenetetrazol hydrochloride reduced M3 receptor levels in the model group, resulting in a significant reduction in primary dysmenorrhea.
Cuscuta reflexa (MECR) extract exhibited a strong binding affinity to 5-hydroxytryptamine receptors, COX-1, COX-2, and M3 receptors through molecular docking. Animal experiments have shown that MECR can significantly reduce the number of writhing episodes induced by acetic acid and the number of pain responses (licking) induced by formalin in mice (171).
In summary, M3 receptors are closely linked to pain management and modulation. Targeting these receptors may lead to innovative approaches to pain management.
|
|
M3RS' CLINICAL APPLICATION AND DRUG RESISTANCE |
|---|
The Selectivity and Therapeutic Efficacy of Agonists or Antagonists in Therapeutic Drugs or compounds
Developing selective drugs with reduced side effects based on structural design is a challenging task. However, with advancements in pharmaceutical science and the support of molecular docking technology, there are promising prospects in this area. Ligands with diverse selectivity offer the potential for various therapeutic effects (see Table 2). Although different drugs targeting the M3 receptor may have distinct action sites and functions, some of them share structural similarities (see Figure 2).
|
|
The latest research shows that an effective technique has been developed to capture M3 receptor ligand binding interactions and distinguish between M3 receptor agonists and antagonists. This technology utilizes a combination of conformation-specific peptide BJ-PRO-13a and the HaloTag trap system. This innovation has improved the efficiency of receptor chromatography in determining binding affinity in new drug development, providing prospects for screening and characterizing active compounds, especially in complex natural products (172).
M3Rs play a crucial role in modulating cell proliferation, differentiation, migration, and adhesion through various physiological reactions in the body. Their clinical significance is vast, as they represent potential therapeutic targets for numerous psychiatric and neurological diseases, including Alzheimer's disease (AD), schizophrenia, Huntington's disease (HD), as well as common internal diseases like type Ⅱ diabetes and chronic obstructive pulmonary disease (COPD). Research on ligands that interact with these receptors holds promise for developing novel pharmaceutical mechanisms to treat neuropsychiatric and neurological disorders effectively. Muscarinic acetylcholine receptor ligands can be categorized into non-selective and selective ligands based on their affinity for specific receptor subtypes. While non-selective ligands have practical utility and are commonly used, selective ligands offer greater potential and scientific research opportunities in the field of precise drug delivery. Selective ligands with partial or absolute specificity for a specific receptor subtype enable the design of drugs targeting receptors activated by endogenous neurotransmitters in specific tissues or organs. This holds significant clinical importance for the development of new and more effective treatments. However, achieving absolute selectivity remains challenging due to the distinctiveness between receptor subtypes. Another reason for limited selectivity may be the common use of traditional pharmaceuticals in large doses, making it difficult to precisely target the disease site.
Multi-Target Drug Delivery Related to M3Rs
Emphasizing the identification of key molecular targets located near lesions is crucial, as it aids in constructing a more comprehensive biological network model and clarifying the synergistic effects of multi-target drugs (214).
Originally, single-target agents (STAs) were considered "clean" drugs with high selectivity, but in practice, their selectivity was often lower than expected, leading to limited effectiveness for different individuals (215). Certain diseases with complex pathogenesis cannot be adequately addressed by the monogeneity of drug molecular targets embodied by STAs. Inspired by these challenges, researchers propose designing multi-target agents (MTAs) that can activate multiple receptors to produce robust and long-lasting efficacy. This approach resembles the concept of "me-too" drugs, which achieved success through molecular modification and structural transformation (216), and contributes to an exhaustive receptor-ligand theory in drug therapy (217, 218).
MTAs have been regarded as "dirty" drugs due to safety concerns and potential side effects, but they are more suitable for diseases with multiple, repeated heteropathogenesis patterns. MTAs trigger the activation of multiple binding sites, resulting in stronger pharmacological effects. Compared to traditional drug delivery methods, synergistic effects produced by modulating multiple targets can achieve biological effects at lower blood concentrations, reducing adverse drug reactions (ADRs) and accumulated toxicity. Moreover, MTAs can prevent receptor desensitization and resistance, maintaining efficacy and improving clinical outcomes. For example, anti-inflammatory drugs targeting histamine H1 receptors, cannabinoid CB2 receptors, muscarinic M3 receptors, and epinephrine β2 receptors produce activating effects, ensuring that inhibition of one receptor does not diminish the overall anti-inflammatory effect (219).
While MTAs offer advantages, they are not without drawbacks. Recent literature reports that MTAs may cause additional ADRs. For instance, antipsychotic drugs with multiple targets in the central nervous system can lead to severe side effects and reduced patient compliance. Despite the challenges, the innovative progress from single-target to multi-target drugs holds promise for individualized drug delivery and optimized treatment. This development can expand clinical and mechanistic knowledge, inspire creative structure modifications, and drive the advancement of new drugs in the future. Future pharmaceutical research may focus on a "dial in and dial out" strategy to improve clinical effects by designing agents that avoid capturing targets that may cause side effects. The multi-target feature of a single compound can be applied to targeted therapy for complex diseases with dose-dependent and time-dependent features.
|
|
CONCLUSION AND PROSPECT |
|---|
The Muscarine Acetylcholine M3 receptor of the GPCR superfamily plays a crucial role in mediating biochemical and physiological reactions through the PLC-DAG-PKC and PLC-IP3-Ca pathways. M3 receptors present attractive therapeutic targets in clinical practice, and different agonists and antagonists of M3 receptors have significant pharmacological effects in pathological situations, including the treatment of opioid addiction. The design of multi-target drugs, encompassing M3 receptors, offers the potential for creating medications that activate multiple receptors to produce powerful and long-lasting effects, overcoming the limitations of single-target drugs with high selectivity but easy resistance. However, further research is needed to fully understand the impressive benefits of M3Rs, which continue to show profound therapeutic effects in clinical practice.
|
|
AUTHOR CONTRIBUTIONS |
|---|
Investigation, S.C.; writing, S.C.; editing, J.W. All authors have read and agreed to the published version of the manuscript.
|
|
INSTITUTIONAL REVIEW BOARD STATEMENT |
|---|
Not applicable.
|
|
INFORMED CONSENT STATEMENT |
|---|
Not applicable.
|
|
DATA AVAILABILITY STATEMENT |
|---|
Not applicable.
|
|
CONFLICTS OF INTEREST |
|---|
The authors declare no conflict of interest.
|
|
REFERENCES |
|---|