| ORIGINAL ARTICLE |
|
|
1 Research & Development Division, Oryza Oil & Fat Chemical Co., Ltd., 1 Numata, Kitagata-cho, Ichinomiya, Aichi 493-8001, Japan;
2 Pias Co. Ltd., 1-3-1 Murotani, Nishi-ku, Kobe 651-2241, Japan
Corresponding Author: Hiroshi Shimoda, Research & Development Division, Oryza Oil & Fat Chemical Co., Ltd., 1 Numata, Kitagata-cho, Ichinomiya, Aichi 493-8001, Japan. Tel: +81-586-86-5141; Fax: +81-586-86-6191; E-mail: kaihatsu@mri.biglobe.ne.jp.
Running title: Tiliroside enhances hyaluronan synthase
| |
ABSTRACT |
| INTRODUCTION | |
|
|
MATERIALS AND METHODS |
|
|
RESULTS |
|
|
DISCUSSION |
|
|
CONCLUSION |
|
|
CONFLICT OF INTEREST |
|
|
AUTHORS' CONTRIBUTIONS |
|
|
FUNDING |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Strawberry seeds are consumed with the sarcocarps, but their effects on the skin have not been well evaluated. We examined the effect of strawberry seed extract (SSE) and tiliroside, a major flavonol glycoside in SSE, on hyaluronan synthase (HAS) 2 expression and hyaluronan production in mouse skin and human fibroblasts. SSE and tiliroside were given orally to mice on a high-fat diet (HFD) or standard diet, and skin HAS2 mRNA expression and the skin hyaluronan content were evaluated. In addition, HAS2 protein expression and hyaluronan release were determined after fibroblasts were treated with SSE and tiliroside. Furthermore, binding of tiliroside to retinoic acid receptor (RAR) γ was examined. SSE (10 and 50 mg/kg) significantly increased HAS2 mRNA expression and the hyaluronan level in the skin of HFD-fed mice. Tiliroside (0.2 and 1 mg/kg) also increased skin HAS2 mRNA expression in HFD-fed mice. Moreover, SSE (10 mg/kg) increased HAS2 mRNA expression in mice on the standard diet. In human fibroblasts, both SSE (1 μg/mL) and tiliroside (0.3 μg/mL) significantly increased hyaluronan production, with HAS2 protein expression also being increased by SSE (10 μg/mL) and tiliroside (0.1 μg/mL). In the RARγ ligand assay, tiliroside (1 and 10 μg/mL) showed binding to RARγ. In conclusion, SSE and tiliroside increased hyaluronan synthase 2 expression in mouse skin and binding of tiliroside to RARγ was suggested to be involved.
KEY WORDS: strawberry; tiliroside; hyaluronan; fibroblast; hyaluronan synthase; retinoic acid receptor|
|
INTRODUCTION |
|---|
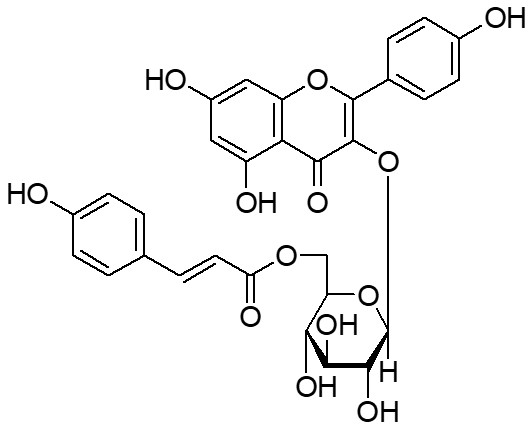
Strawberries (Fragaria × ananassa Duch., rosaceae) are globally cultivated. The fruit is rich in polyphenols (1-3) for which many biological functions have been reported, including anticarcinogenic (4, 5), vasodilatory (6), antidiabetic (7), antithrombotic (8), photoprotective (9) and neuroprotective (10) activities. However, few scientific studies of the seeds are available despite the fact that the seeds are also eaten. Tsukamoto et al. (11) reported that tiliroside (Fig. 1), a flavonol glycoside, is contained in strawberry seeds. Tiliroside is also found in other edible parts of plants, such as linden leaves (12) and rosehip fruits (13), and has been reported to exhibit hepatoprotective (12), antiobesity (13), antidiabetic (14), antioxidant (15), anti-inflammatory (16) and antimelanogenic (17) effects. We recently reported that an aqueous ethanol extract of strawberry seeds (SSE) and tiliroside promote production of ceramides in the skin, contributing to moisturization of the epidermis (18).
Hyaluronan is another skin moisturizer, which is produced by dermal fibroblasts and released into the extracellular matrix (19). Hyaluronan consists of glycosaminoglycans containing hexosamine and contributes to both moisture retention and elasticity of the dermis (20). Synthesis of hyaluronan is regulated by hyaluronan synthase (HAS) 1-3, with HAS2 playing a crucial role in dermal hyaluronan synthesis (21). Several pathways are involved in regulating HAS2 expression, such as the transforming growth factor (TGF)-β/Smad pathway (22), cAMP/cAMP-response element-binding protein (CREB) pathway (22), and platelet-derived growth factor (PDGF) pathway (23). Retinoic acid (RA) enhances hyaluronan production and is well known to increase HAS2 expression (24). Binding of RA to the RA receptor (RAR) activates the cAMP/CREB pathway (25), leading to upregulation of HAS2 mRNA expression (26), suggesting that RAR ligands could promote hyaluronan production via HAS. Tiliroside has been reported to upregulate peroxisome proliferator-activated receptor (PPAR)α expression (13). Since PPARα forms heterodimers with RAR, tiliroside may promote hyaluronan production via the PPARα/RAR pathway. Against this background, we evaluated the effects of SSE and tiliroside on HAS2 expression in mouse skin and human fibroblasts. We also assessed binding of tiliroside to the RAR to investigate the mechanism of its influence on HAS2 expression.
|
|
MATERIALS AND METHODS |
|---|
Preparation of SSE and measurement of the tiliroside content
Strawberry seeds cultivated in California were purchased from Premier Specialties, Inc. (Middlesex, NJ, USA). The seeds (175 g) were powdered and defatted twice with n-hexane (400 mL), after which the residue was extracted with 500 mL of 70% (v/v) ethanol for 2 h at 80°C. The solvent was subsequently removed by evaporation to obtain SSE (yield: 2.3%) Then the tiliroside content of SSE was determined by HPLC, which was performed by using a C18 column (250 × 4.6 mm i.d.) (Capcell Pak C18, Shiseido Co., Ltd., Tokyo, Japan) at a flow rate of 1 mL/min with 35% acetonitrile as the solvent UV detection was performed at a wavelength 254 nm and tiliroside was identified by direct comparison with authentic tiliroside (Nagara Science Co. Ltd., Gifu, Japan). HPLC revealed that the tiliroside content of SSE was 2.2%.
Reagents
An RNAeasyTM Protect Mini Kit was purchased from Qiagen (Hilden, Germany). Dulbecco’s modified Eagle’s medium (D-MEM), penicillin-streptomycin mixture, and 3,3’-diaminobenzidine (DAB) were obtained from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Fetal calf serum (FCS), random hexamers, 10 mM dNTP mixture (PCR grade) and an RNase inhibitor were sourced from Invitrogen Co. (Carlsbad, CA, USA). PrimeScriptTM Reverse Transcriptase and SYBR ® Green I were purchased from Takara Bio Inc. (Otsu, Japan). QnE Hyaluronic Acid (HA) ELISA Assay was obtained from Biotech Trading Partners (Encinitas, CA, USA). Mouse anti-hyaluronan binding protein (HBP) 2 monoclonal antibody (1HA) was purchased from Abnova (Taipei, Taiwan), rabbit HRP-conjugated anti-mouse immunoglobulin and CSA II were from DAKO (Glostrup, Denmark), and goat anti-HAS2 polyclonal IgG (S-15: sc-34067) was from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). In addition, HRP-conjugated rabbit anti-goat IgG was obtained from MP Biochemical, LLC (Solon, OH, USA), rabbit anti-GAPDH polyclonal antibody was from Imgenex (San Diego, CA, USA), and HRP-conjugated goat anti-rabbit IgG was from Millipore (Billerica, MA, USA). EnBio RCAS for PPAR (alpha) was purchased from EnBioTec Laboratories Co. Ltd. (Tokyo, Japan) and NuLigand RARγ was obtained from Micro Systems (Kyoto, Japan).
Animals and cells
Male ddY mice (10 to 11 weeks old) and female hairless mice (Hos: HR-1, 5 weeks old) were obtained from Japan SLC, Inc. (Shizuoka, Japan) and Hoshino Laboratory Animal Inc. (Saitama, Japan), respectively. The mice were housed in an air-conditioned room (23 ± 1°C, 50 ± 10% R.H.) for at least 3 days and were allowed free access to a standard non-purified diet (CE-2, Clea Japan Inc., Shizuoka, Japan) and tap water. High-fat diet (HFD) 32 was obtained from Clea Japan Inc. (Tokyo, Japan). It has a 32% fat content, consisting of oleate (64.3%), palmitate (12.6%), linolate (3.26%), and triglycerides of other free fatty acids. All experiments were performed in accordance with the Guidelines for the Proper Conduct of Animal Experiments (Special Council of Japan, June 1, 2006). These experiments were approved by the ethical committee of the research and development section of Oryza Oil & Fat Chemical Co. Ltd. on December 7, 2007, July 31, 2008, April 1, 2009, May 1, 2009, June 19, 2009, and September 23, 2009 (5 certification sheets). The animal study was performed before September 1, 2008. NB1RGB, a human fibroblast cell line derived from a Japanese neonate, was obtained from Riken Bioresources Bank (Tsukuba, Japan).
Treatment of mice with SSE and tiliroside
In the initial experiment, SSE (10 and 50 mg/kg) or tiliroside (0.2 and 1 mg/kg) was given orally for 8 days to ddY mice on the HFD. Normal control mice received CE-2 (standard diet) instead of the HFD. SSE (10 and 50 mg/kg) suspended in 5% gum Arabic in water was administered orally once a day. On the day 7, the mice were fasted for 18 h and anesthetized with ether, after which a skin specimen was removed from the middle of the back. Each skin specimen (approximately 200 mg) was soaked in RNA laterTM from an RNAeasyTM Protect Mini Kit and was stored at 4°C until evaluation of HAS2 mRNA. In the second experiment, SSE (10 and 50 mg/kg) was given orally for 8 days to hairless mice on the HFD or standard diet. After fasting for 18 h, mice were anesthetized and a skin specimen was collected for determination of HAS2 mRNA expression and for immunohistochemical staining to detect hyaluronan binding protein 2 (HBP2). The specimen for immunostaining was soaked in 4% paraformaldehyde and stored at 4°C.
Cell culture and measurement of hyaluronan
NB1RGB cells (5×104/mL) were suspended in D-MEM containing 10% FCS, 100 units/mL penicillin, and 100 μg/mL streptomycin. The cell suspension (500 μL) was seeded into a 24-well microplate for evaluation of hyaluronan production. After culture for 3 days, the culture medium was collected from the plate for determination of hyaluronan using the QnE Hyaluronic Acid (HA) ELISA Assay. For western blotting of HAS2 protein, 10 mL of cell suspension (3 × 104/mL) was seeded into a φ 14 cm dish. After incubation for 24 h, SSE or tiliroside dissolved in 1% DMSO was added to the dish (final DMSO concentration: 0.1%). Before western blotting, the medium was removed from the dish and the cells were washed with warmed phosphate-buffered saline (PBS). Then the cells were harvested with 1 mL of ice-cold lysis buffer (50 mM Tris-HCl, 150 mM NaCl, and 1% Triton X-100; pH: 7.2) and were stored at -20°C.
Real-time RT-PCR analysis
Total RNA was extracted and purified from stored cell lysates by using the RNAeasy TM Protect Mini Kit, after which cDNA was synthesized using random hexamers, dNTP mixture, PrimeScriptTM Reverse Transcriptase, and an RNase inhibitor. Real-time RT-PCR was performed with a Thermal Cycler DiceTM Real-Time System (TM800, Takara Bio Inc.) using SYBR® Green I and primers (5′→3′) for Has2 (forward: GTCCAAGTGCCTTACTGAAACTCCC, reverse: GAGGATGTTCCAGATTTTACCCCTG) or β-actin (forward: aatcgtgcgtgacatcaaag, reverse: gaaaagagcctcagggcat).
Immunohistochemical staining of HBP in mouse skin
Paraffin sections were produced by using a Young-type sliding microtome (Sakura Finetek Japan Co. Ltd., Tokyo, Japan) and a disposable microtome blade. Paraffin-embedded specimens of mouse skin were deparaffinized and activated by autoclaving (120°C, 10 min). Then the specimens were treated with 1% H2O2 in methanol, blocked with skim milk, and incubated overnight at 4°C with anti-HBP2 antibody (1:100 dilution). After washing with Tris-buffered saline (TBS), the specimens were incubated with undiluted HRP-conjugated anti-mouse immunoglobulin solution for 15 min. After washing the specimens again, signals were amplified with the CSA II system, followed by nuclear staining with Mayer’s hematoxylin-eosin (HE). DAB and HE staining were observed under a microscope.
Western blotting
Cell lysate (1 mg protein/mL) was mixed with an equal volume of Laemmli sample buffer [62.5 mM Tris-HCl, 2% sodium dodecyl sulfate (SDS), 5% 2-mercaptoethanol, 25% glycerol and 0.01% bromophenol blue] and heated at 95°C for 5 min. Then an aliquot of the sample solution (20 μL) was subjected to electrophoresis on 10% SDS gel. Proteins were transferred to a polyvinylidene fluoride membrane, which was probed for HAS2 by using goat anti-HAS2 (0.4 μg/mL, 1:500 dilution) antibody and HRP-conjugated anti-goat IgG (23.7 μg/mL, 1:1,000 dilution), with DAB and H2O2 for detection. For identification of GAPDH, rabbit anti-GAPDH (1:5,000 dilution) antibody and HRP-conjugated anti-rabbit IgG (1:12,500 dilution) were used. Detection was performed by the chemiluminescence method with an ECL Plus Blotting Detection system and Amersham HyperfilmTM ECL.
Ligand assay for PPARα and RARγ
The activity of tiliroside as a ligand for PPARα or RARγ was evaluated by using commercial kits [EnBio RCAS for PPAR (alpha) and NuLigand for RARγ] according to the manufacturer’s instructions.
Statistical analysis
Results are expressed as the mean and S.E. Significance of differences was examined by one-way ANOVA, followed by Dunnett’s test, and p<0.05 was considered to indicate significance.
|
|
RESULTS |
|---|
Effect of SSE and tiliroside on skin expression of hyaluronan and HAS2 mRNA in HFD mice and standard diet mice
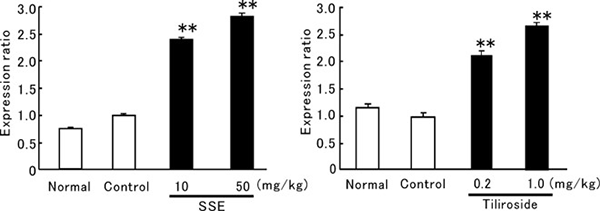
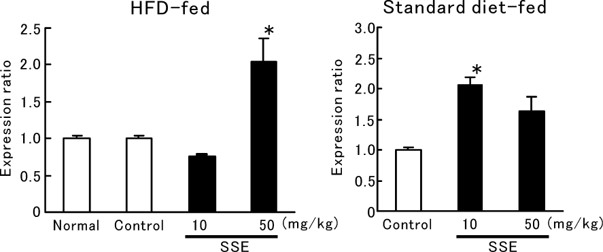
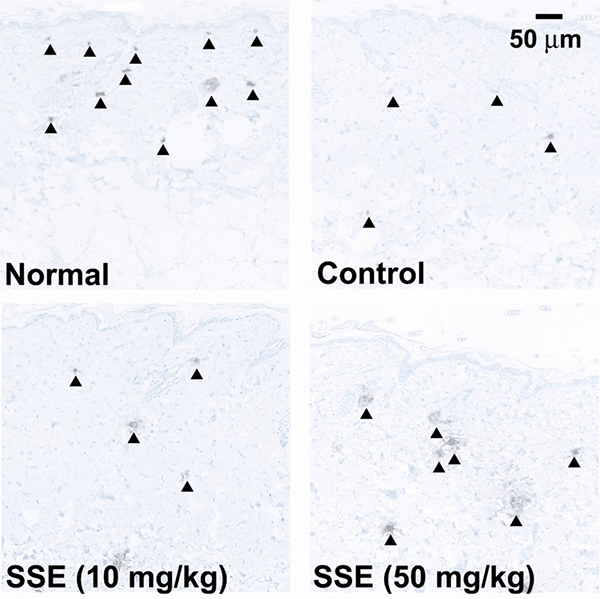
The effect of SSE on skin expression of HAS2 mRNA was examined in ddY mice fed the HFD. As shown in Fig. 2A, HAS2 mRNA expression was significantly increased by oral administration of SSE (10 and 50 mg/kg) for 8 days. Tiliroside also clearly upregulated HAS2 mRNA expression at 0.2 and 1 mg/kg (Fig. 2B). Similar results were obtained in hairless mice fed the HFD and administered SSE (50 mg/kg) for 8 days (Fig. 3A). Moreover, SSE (10 mg/kg) significantly enhanced HAS2 mRNA expression in hairless mice on the standard diet (Fig. 3B). Figure 4 shows images of skin sections from mice that were fed the HFD and administered SSE. HBP2 is stained brown (arrowhead). The stained area was decreased in the skin of HFD mice (Fig. 4B), while the stained area was increased in the skin of mice given SSE (10 and 50 mg/kg) (Fig. 4C, 4D).
Hyaluronan production and HAS2 protein expression by human fibroblasts
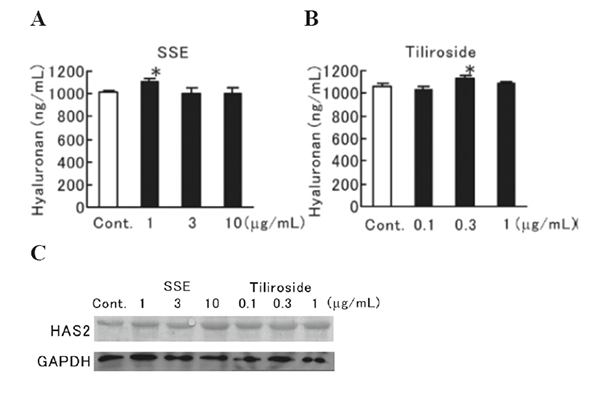
Hyaluronan production and HAS2 protein expression were evaluated in human fibroblasts treated with SSE or tiliroside. SSE (1 μg/mL) significantly enhanced hyaluronan production (Fig. 5A), and tiliroside (0.3 μg/mL) also significantly increased hyaluronan production (Fig. 5B). In addition, SSE (10 μg/mL) and tiliroside (0.1-1 μg/mL) both clearly increased HAS2 protein expression (Fig. 5C).
Binding of tiliroside to PPARα and RARγ
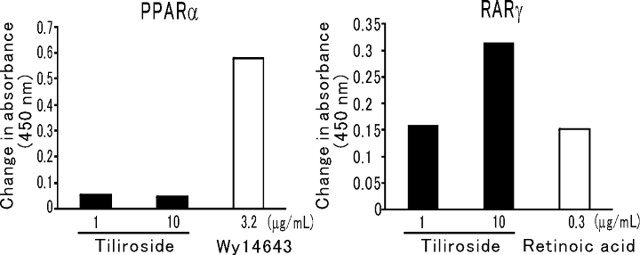
This assay did not detect an increase of absorbance indicating the binding of tiliroside to PPARα (Fig. 6A). On the other hand, there was an increase of absorbance indicating the binding of tiliroside (1 and 10 μg/mL) to RARγ (Fig. 6B). Tiliroside was suggested to have approximately one-third of the affinity of RA for RARγ.
|
|
|
|
|
|
|
|
DISCUSSION |
|---|
The morphological changes of skin architecture accompanied by accumulation of subcutaneous fat in obesity have not been well established. An increase of subcutaneous adipose tissue impairs dermal elasticity (27) and also shows a negative correlation with the dermal collagen content (28). In rats with dietary obesity, reduction of the dermal content of collagen and hyaluronan has been confirmed (29). It was also reported that an increase of adipose tissue is followed by a decrease of dermal fibroblasts in obese mice (30). These findings suggest that accumulation of subcutaneous fat affects the dermal architecture and alters the composition of the extracellular matrix.
Tiliroside in SSE previously demonstrated an antiobesity effect along with enhancement of PPARα expression (13). Therefore, we evaluated the antiobesity effects of SSE and tiliroside by examining the skin of mice fed an HFD. We found that SSE caused significant upregulation of HAS2 mRNA expression in the skin and also increased hyaluronan mRNA expression. Enhancement of HAS2 mRNA expression by SSE was also seen in mice on a normal diet. Oral administration of tiliroside increased HAS2 mRNA expression in the skin at a lower dose than that of SSE. The intestinal absorption mechanism of tiliroside (31) and its effects on lipid membrane (32) have been studied in detail, and these mechanisms may help tiliroside to exhibit activity at low doses. Intrinsic factors and hormones that enhance HAS2 expression have been reported such as adiponectin (33), collagen fragments (34), cyclic AMP (22), IL-1α (35), and TNF-α (35). On the other hand, steroids (36), ultraviolet B light (37) and oxidative stress (38) suppress HAS2 expression. Tiliroside is a low molecular weight polyphenolic constituent with antioxidant activity (39), so its structure is quite different from the peptides and glycoproteins mentioned above. Accordingly, a different mechanism may be involved in the effect of tiliroside on HAS2 mRNA expression.
We investigated the direct effects of SSE and tiliroside on hyaluronan production and HAS2 protein expression by cultured human fibroblasts. As a result, SSE only caused a slight increment of hyaluronan release into the culture medium from fibroblasts, although HAS2 protein expression was clearly enhanced. Tiliroside also slightly increased hyaluronan production and clearly upregulated HAS2 protein. There have been no reports about low molecular weight compounds that enhance HAS2 expression, except for compounds derived from musk oil, including ethylene brassylate, astratone, and 1,4-dioxacycloheptadecane-5,17-dione (called musk T) that promote HAS expression by binding to PPARα (40). Tiliroside was previously reported to increase hepatic expression of PPARα in mice on an HFD (13), while RA upregulates hyaluronan production in fibroblasts (41), mouse skin (42), and human skin (43). Binding of RA to the RAR is the primary initiator of hyaluronan production (25, 26). Accordingly, we evaluated binding of tiliroside to PPARα and RARγ in this study. While affinity for PPARα was not confirmed, tiliroside exhibited binding to RARγ at both 1 and 10 μg/mL. Therefore, binding of tiliroside to RARγ may be involved in the upregulation of HAS2 expression by SSE.
|
|
CONCLUSION |
|---|
SSE and tiliroside (a flavonoid component of SSE) caused an increase of HAS2 mRNA expression in mouse skin and human fibroblasts. Tiliroside showed binding to RARγ . These findings suggest that hyaluronan production was promoted via binding of tiliroside to RARγ and subsequent upregulation of HAS2 expression.
|
|
CONFLICT OF INTEREST |
|---|
Junji Tanaka Ph. D. and Hiroshi Shimoda Ph.D. are employed by Oryza Oil & Fat Chemical Co. Ltd. as research scientists. Oryza Oil & Fat Chemical is the developer and manufacturer of the strawberry seed extract used in this study. The authors have not received personal financial gain from the sales of this product. All findings and views expressed in this paper are those of authors and do not necessarily reflect the view of Oryza Oil & Fat Chemical. Kaori Zaiki and Koichi Nakaoji are employees of Pias Co. Ltd. The authors have not received personal financial gain from sales of this product. All findings and views expressed in this paper are those of authors and do not necessarily reflect the view of Pias Co. Ltd.
|
|
AUTHORS' CONTRIBUTIONS |
|---|
Hiroshi Shimoda Ph. D. and Junji Tanaka Ph. D. conducted the study and performed the tests, except for the PPARα and RARγ assays, and wrote the manuscript. Kaori Zaiki and Koichi Nakaoji performed the PPARα and RARγ assays.
|
|
FUNDING |
|---|
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sector.
|
|
REFERENCES |
|---|