| Original Article |
|
|
1 Monash University, School of Biological Sciences, Victoria 3800, Australia;
2 St. Mary’s College, University of Melbourne, P.O. Box 140, Parkville, Victoria 3052, Australia;
3 Department of Molecular Biology, University of Central Florida, Orlando Fl, 32826, USA;
4 International Center for Genetic Engineering and Biotechnology, Aruna Asaf Ali Marg, New Delhi 110067, India
Corresponding author: Jamiu A. Ogunbanwo, Monash University, School of Biological Sciences, Victoria 3800, Australia. Tel:+61-3-9905 9464; Fax:+61-3-93491335; E-mail:jamiu.ogunbanwo@sci.monash.edu.au. Note: The data from this paper were presented at the MIM Pan-African Malaria Conference in Arusha, Tanzania in 2002 and XI International Congress of Parasitology in 2006, Glasgow, Scotland-UK.
Short title - page header: expression, purification and Characterization of Plasmodium vivax trap
| |
ABSTRACT |
| INTRODUCTION | |
|
|
MATERIALAS AND METHODS |
|
|
RESULTS AND DISCUSSIONS |
|
|
ACKNOWLEDGEMENT |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Thrombospondin Related Adhesive Protein (TRAP) is a transmembrane parasite molecule responsible in sporozoite-host interactions. This molecule is one of the most promising vaccine candidates against the pre-erythrocytic forms of malaria. In the present study, a gene encoding the Plasmodium vivax TRAP (PvTRAP) was expressed in Escherichia coli (M15 strain) using the expression plasmid pQE30. The expressed recombinant
protein PvTRAP of about 70kDa was achieved, purified and refolded according to the standardized refolding procedure. This refolded protein (PvTRAP) showed a single band monomeric form with SDS-PAGE and blot analysis. In reduced and alkylated form, PvTRAP showed less binding to hepatoma (HepG2) liver cells, when compared to the normal purified and refolded form. Purified and refolded recombinant PvTRAP bound Duffy-positive human erythrocytes, while no binding was observed with Duffy-negative erythrocytes. Our report on PvTRAP is currently documented for the first time and it has been able to provide an experimental evidence of the biochemical and binding properties of PvTRAP in the invasion of hepatocytes and interaction with Duffy-positive and Duffy-negative human erythrocytes. In conclusion, our findings have been able to demonstrate the potential of PvTRAP as a promising target for vivax malaria vaccine candidate.
KEY WORDS:
thrombospondin-related adhesive protein (TRAP); plasmodium vivax; malaria
|
|
INTRODUCTION |
|---|
The reported global human malaria stands at 300-500 million clinical cases with an average of about 2 million deaths per year (1a). Plasmodium falciparum causes 80% of human malaria’s morbidity and mortality, mostly in sub-Saharan Africa, however, Plasmodium vivax annually accounts for 70-80 million cases across much of the tropics and subtropics of the world (1b) Plasmodium vivax is the second most prevalent species and widely distributed human parasite that causes malaria morbidity among people of all ages in Africa, Asia, the Middle East and Latin America (1c). Several antigens expressed at different stages of the parasite life cycle have been characterized and found to have the potential for use in a sub unit vaccine against P. vivax (2-5, 43-45).
One of the antigens thrombospondin-related adhesive protein (TRAP), a potential malaria vaccine candidate, is one of the two major proteins identified on the sporozoite surface of Plasmodium species that are involved in hepatocyte or HepG2 cell line recognition / or invasion (6-11). The genes encoding TRAP and the circumsporozoite TRAP related protein (CTRP) are differentially expressed in sporozoites and ookinetes, respectively, two motile forms of Plasmodium species found exclusively during the life cycle of the parasite in the mosquito (6, 12-14).
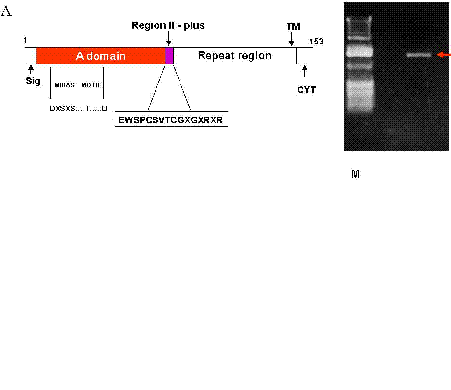
TRAP is found in the micronemes and a type 1 transmembrane protein (Fig 1) whose ectodomain consists of (i) an A domain, (ii) a TSR, and (iii) a repeat region of variable length and sequence, depending on the plasmodial species. The A domain is a ~ 200 residue -long adhesive module that was first recognized in the plasma protein von Willebrand factor (15). It now defines a superfamily of soluble proteins, including complement protein factor B and C2, extra cellular matrix proteins, including numerous types of non fibrillar and FACIT (fibril associated collagen with interrupted triple helix) collagens, and integral membrane proteins, including seven integrin and chains (6, 15).
The crystal structures of the A domains of TRAP have been determined (16-19). They comprise alternating amphiphatic and helices and hydrophobic B strands conforming to the classic and B “Rossmann” fold (15). Identification of 5 residues which coordinate a divalent cation (Mg2+ or Mn2+) in A domains, define the metal-ion dependent adhesion site (MIDAS) motif (Fig 1a), which is conserved in a number of A domains of TRAP (15). Mutational analysis of the metal ion-dependent adhesion site motif in integrin-a chain has demonstrated the critical role of these five residues in ligand-binding (20-24).
In human living in malaria endemic areas, reports have shown close correlation between levels of TRAP antibodies and clinical protection against malaria (25). Also, a Babesia bovis merozoite protein was discovered to have domain architecture highly homologous to Plasmodium falciparum TRAP (45). Antisera to the TSR region of TRAP inhibits the in vitro invasion of erythrocytes by the asexual blood stage merozoites and also recognizes a TRAP-like protein in the blood stage lysate of P. falciparum (26). Therefore the possible role of TRAP in two most important different stages of the parasite: the sporozoite invasion of hepatocytes and merozoite invasion of erythrocytes makes the molecule a potential malaria vaccine candidate.
We have expressed PvTRAP nearly full length gene in Escherichia coli, purified and refolded the protein with established standard procedures and characterized the protein to ascertain its biological and functional activities as a potential target for the development of a protective and safe vivax malaria vaccine.
|
|
MATERIALS AND METHODS
|
|---|
Amplification and Cloning Strategy for PvTRAP gene. We obtained blood from an Indian P. vivax infected patient by venipuncture and passed through CF-11 column (Whatman) to remove leukocytes. The parasitized erythrocytes were purified by centrifugation on a ficoll-hypaque gradient and subjected to saponin lysis to get a sporozoite rich preparation. DNA was isolated from the sporozoite rich preparation by standard procedures (27).Based on sequence comparison of PvTRAP with PkTRAP and PgTRAP (9) a set of primers from the conserved sequences, one near the putative signal peptide cleavage site, forward primer.
(5’-GCGGATCCGACGAAATAAAGTATAGTGAAGAAGTATG-3’) and the other at the extreme carboxyl-terminus,a reverse primer (5’-CACTCAAGCTTAAATTTTGTA GCCATTATT-3’) were synthesized.
PCR amplification was achieved using 100 ng of P. vivax genomic DNA in steps, with initial hot start of 94ºC/3 min followed by 5 cycles of 94ºC/1 min, 44ºC/2 min and 72ºC/3 min and 25 cycles of 94ºC/50s, 48ºC/1 min and 72ºC/3 min.
A fragment of about 1.4 Kb was amplified from P. vivax genomic DNA. The PCR product was gel purified using QIA quick gel extraction kit (QIAGEN) and cloned into the pGEM-T cloning vector (Promega) as per the manufacturer’s instructions.
Positive clones were selected by Southern hybridization and restriction analysis sequencing was performed using the dideaxy chain termination method (Sequenase, USB), with vector specific and gene specific sequencing primers. Eight independent clones were sequenced to rule out the possibility of any PCR generated artifacts. Sequence analysis and alignment were carried out using MAC-VECTOR and DNASTAR (data not shown).
Expression, purification and refolding of recombinant PvTRAP. The PCR fragment of about 1.4 Kb full lengths PvTRAP gene was digested with Bam HI and Hind III and then sub-cloned into pQE-30 plasmid vector which had been digested with the same restriction enzymes. The resulting ligation product- pQE-30-His tagged PvTRAP was used to transform the E. coli (M15) cells. Ampicillin and Kanamycin resistant clones containing pQE30-His-6-PvTRAP plasmid were identified by restriction with Bam H1 and Hind 111 (Fig 1b). Competent E. coli M15 cells were transformed with plasmid pQE-30-His-6-PvTRAP and transformants were selected on LB medium containing Ampicillin (100µg/ml) and Kanamycin (25µg/ml). A single colony was inoculated into 50 ml of LB medium containing the same antibiotics and grown at 37ºC; 200 r.p.m. until OD600 reached about 0.6-0.7. An aliquot (5ml) of this pre-culture was transferred into 1 liter LB medium containing the same antibiotics, incubated on a shaker at 37ºC; 200 r.p.m. for 2-4 hr. The protein expression was induced by addition of isopropyl-β�����������������-D-thiogalactopyranoside (IPTG) to a final concentration of 1mM when OD600 reached 0.6-0.7.
After a 6-8 hr post incubation period, the bacterial cells were harvested by centrifugation at 5000g at 4ºC for 45 min. The induced and uninduced expressed PvTRAP was comparatively analyzed on SDS-PAGE (data not shown). The cell pellet obtained after harvesting the bacterial cells (E. coli) was suspended in ice-cold TBS (20mMTris-Cl, pH 7.5, 250mM NaCl).
The washed E. coli cell pellet containing PvTRAP (insoluble protein in inclusion bodies) was thawed in ice and solubilized in lysis buffer (6MGu.HCl, 20mMTris-Cl, PH 8.0, 250 mM NaCl; 5mL/g wet weight of the pellet) at room temperature for 60 minutes. After centrifugation (10,000xg 30 min. 4ºC), the supernatant was incubated with Nickel-Nitriloacetate acid (Ni-NTA) agarose resin (1.0 ml of 50% slurry was used for the lysate from 2 g of the induced bacteria cell pellet) which was already equilibrated with the lysis buffer for 1 hr at room temperature.
The resin-lysate suspension was poured to a Bio-Rad-Poly-Prep Chromatography column (1.5 × 12 cm propylene column) and the unbound proteins were allowed to pass down. Nickel- Nitriloacetate acid (Ni-NTA) agarose resin with bound proteins was washed with (a) lysis buffer (20 × bed volume), (b) 10 × bed volume of wash buffer 1 (8M Urea, 20mM Tris-Cl, ph 8.0, 250 mM NaCl ) and (c) 10 × bed volume of wash buffer 2 (6M Urea, 20 mMTris-Cl, pH 8.0, 250 mM NaCl).
After washing, the bound PvTRAP was eluted successfully with 100 mM imidazole in 6M Urea, 20mM Tris-Cl, pH 8.0.
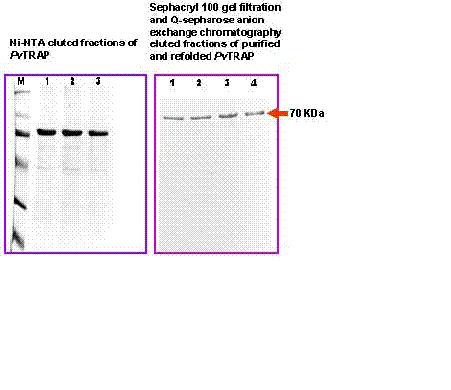
The eluted PvTRAP was further purified by anion exchange chromatography (Q-Sepharose; Pharmacia) under denaturing condition. The anion exchange chromatography was performed by using Fast Performance Liquid Chromatography (FPLC) system (Pharmacia) (Buffer A: 6M Urea, 20 mM Tris-Cl, ph 8.0; Buffer B: 6M Urea, 20 mMTris-Cl, pH 8.0 and 1 M NaCl), at 0.75 ml/min flow rate. Elution of the purified PvTRAP was carried out with increasing concentration of salt (30-40% of Buffer B in about 30 min) and monitored at 280 nm. 1mL elution fractions corresponding to the peak were collected and analyzed by SDS-PAGE for purity. Protein concentration was determined by the method of Bradford (28) using bovine serum albumin as a reference. Protein SDS-polyacrylamide gel was performed by the method described by Chang et al (29). Further purification with Sephacryl 100 (S-100) gel filtration was carried out in order to achieve a single band purified protein (Fig 2). A standardized in vitro refolding procedure was designed and employed for the protein (PvTRAP) with modification of the method described by Glansbeek et al (30).
The purified PvTRAP was rapidly diluted in various buffers containing 20 mM Tris-Cl pH 8.6, 2mM EDTA, 20mM NaCl, 10% Glycerol, 2mM Glutathione (reduced-GSH) and 0.2mM oxidized Glutathione–(GSSG) and the refolding was carried out at the temperature 10ºC within the duration of 40-70 h.
The refolded PvTRAP was concentrated and dialyzed against 200mM Tris pH 7.5, 200 mM NaCl and 1% glycerol at 4ºC for 4 h. The final dialysis was carried out with phosphate buffered saline (PBS) pH 7.2 overnight at 4ºC. The protein (PvTRAP) was refolded immediately after Ni-NTA purification laboratory procedure was carried out.
Characterization of recombinant PvTRAP Circular Dichroism of recombinant PvTRAP. Circular dichroism (CD) spectra of refolded PvTRAP were recorded on a Jasco model J-720 spectropolarimeter at 50-100 µg/ml protein in 10 mM sodium phosphate pH 7.0. All measurements were carried out in a 0.1-mm path length cylindrical cuvette at room temperature at 180-260nm. The number of spectra was recorded at a scan speed of 10nm/min with a step resolution of about 0.1nm (40) (data not shown).
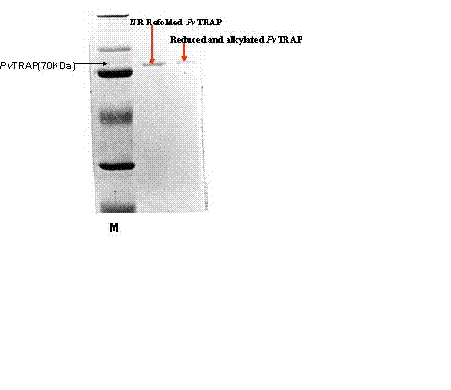
Reduction and Alkylation of recombinant PvTRAP. By modification of the procedure described by Aitkin and Learmonth (the protein biochemistry protocols), the refolded PvTRAP was reduced with DTT (0.02 M final concentration) under N2 for 2hr at 45ºC. The reduced PvTRAP protein was alkylated (in dark) with 0.1 M iodoacetamide with stirring at 37ºC for 1 hr. This reduced and alkylated PvTRAP was desalted on prepared Sephadex 25 column and estimated by Bradford (BioRad) assay accordingly (31).
PvTRAP purified and refolded samples were analyzed by discontinuous SDS-PAGE (29), each sample was denatured in a sample buffer with or without β���������������-Mercapto-ethanol by boiling at 96ºC for 10 min and then used for electrophoresis on 10% SDS/polyacrylamide gel. Protein staining was performed with Coomassie brilliant blue (Fig 3b).
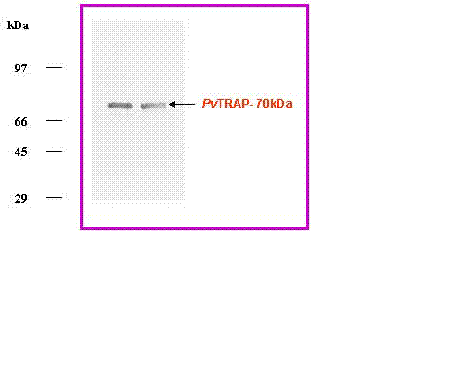
Immunoblotting. For immunologic characterization, anti PvTRAP sera were raised in rabbit and generated in our laboratory for immunoblot assay (32). The purified and refolded PvTRAP was transferred from the unstained SDS gel to a nitro-cellulose system (Biorad) at 990 mA for 2 hr. After blocking with 5% milk in phosphate Buffer Saline PBS containing 0.01% Tween 20 (buffer A), the membrane was incubated with anti-PvTRAP antibodies (1:10,000) raised in rabbit at room temperature for 1 hr. Unbound antibodies were removed by intensive washing in buffer A and incubated with the secondary horse-radish peroxidase conjugate Anti-IgG specific antibody (Sigma) raised in goat. After repeated washing the labeled PvTRAP was detected by colourizing with appropriate substrate – Diaminobenzidine (DAB) (Sigma) for the conjugated secondary antibody (Fig 3a&b). Plasmodium falciparum TRAP (PfTRAP) and Plasmodium cynomolgi (PcTRAP) were used as positive controls to study the cross reactivity with generated anti PvTRAP antibodies (32, 46).
Hepatoma (HepG2) Liver Cells assay. Further characterization of PvTRAP was carried out with HepG2 liver cells in-vitro. The maintenance of HepG2 liver cells (American Type Culture Collection, Rockville, MD, USA) was carried out with Dulbecco Minimum Essential Medium (DMEM) supplemented with 10% Fetal Calf Serum (FCS). The cells were removed from culture flasks with Trypsin-EDTA (0.25% Trypsin, 1mM EDTA in HBSS) (GIBCO-BRL) and pelleted at 3000xg for 15 min at 4ºC. The cell pellet was resuspended in complete DMEM medium to a concentration of 106cells/ml. 105cells/well were plated in 96 well cell culture plates and allowed to grow overnight. The cells were washed with the medium twice at the interval of 10 min and fixed with 4% Para formaldehyde for 15-20 min.
Following washings with TBS (50mM Tris, ph 7.4, 130mM NaCl), the wells were blocked with 1% BSA in Tris buffer saline (TBS) for 2hr at 37ºC. The cells were incubated with different concentrations of the purified and refolded PvTRAP and/or reduced-alkylated purified and refolded PvTRAP for 1 hr at 37ºC.
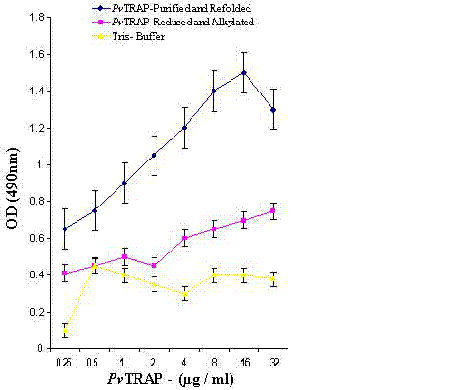
Unbound proteins were removed by washing with TBS three times for 10 min each and incubated with polyclonal antibodies raised in mice against purified and refolded PvTRAP for 45 min at room temperature. The bound antibodies to PvTRAP were detected by goat anti-mouse IgG-horseradish peroxidase (HRP) conjugate (Sigma). Binding of PvTRAP to HepG2 cells was measured at 490 nm using micro plate reader (Molecular Devices–USA). Absorbance obtained at 490 nm was plotted against different concentrations of the PvTRAP using Kaleida graph program (Fig 4).
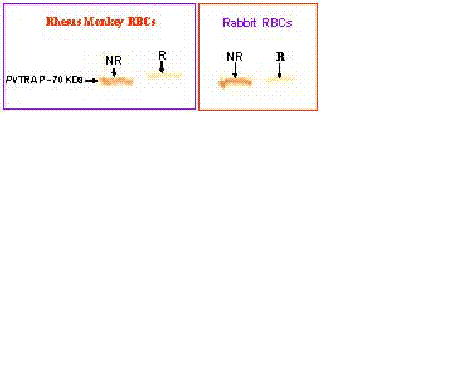
Erythrocyte Interaction Assay. Purified and refolded PvTRAP was further analyzed for its biological activity by employing erythrocyte binding assay according to the established protocols (32, 33). Basically Duffy-positive and Duffy-negative human erythrocytes were washed with incomplete RPMI medium. 0.5 volume of Foetal Complement Serum (FCS) was added to the washed erythrocytes with the corresponding purified and refolded PvTRAP, incubated at 37ºC for 1 hr. The mixture was passed through Dibutylphthalate (DBP) and the supernatant material was eluted with 1.5 M NaCl Western blot analysis was carried out on the eluted supernatant material to confirm the binding of PvTRAP to the erythrocytes using PvTRAP-rabbit raised antibodies to recognize the protein on the blot (Fig 5a&b).
| |
|---|
In the present investigation, expressed and purified PvTRAP of about 30mg/litre recombinant protein was obtained. This showed that the nearly full length PvTRAP gene (~1.4 Kb) that encoded the protein was fully expressed, as shown in Figure 3a-b. Our study revealed that the recombinant PvTRAP was insoluble and present in inclusion bodies. We are reporting for the first time the expression, purification and characterization of PvTRAP, even though, earlier studies on TRAP only reported the complete sequence of P. yoelii sporozoite surface protein 2 (SSP2) gene (34), characterization of P. falciparum SSP2 (6), cloning and cross species comparison of P. knowlesi, P. vivax and P. gallinaceum (9), and also sequence analysis of P. cynomolgi TRAP (10). In the present study, eluted recombinant PvTRAP was subjected to multi-stage purification procedure, as shown in Figure 2. Single band purified, refolded PvTRAP was generated (Fig 3a) and further analyzed under different conditions (Fig 3a&b, 4, 5a-b). Circular dichroism (CD) spectra and helical conformation of PvTRAP carried out using J-720 spectropolarimeter- computer based analytical tools confirmed the conformational position of the protein at 180-260nm. The spectrum of PvTRAP recorded at pH 7.2 showed a minimum at 206nm and a shoulder at about 240nm, typical of �����-helical protein, thus, confirming that the recombinant purified PvTRAP was structurally conformed and refolded (data not shown).
The binding of refolded PvTRAP under different conditions, as shown in Figure 4, fully showed the capacity of this recombinant protein of binding to HepG2 liver cells. This confirmed the biological activity and structural conformation of purified and refolded PvTRAP. These results suggest that P. vivax TRAP (PvTRAP) gene which encoded the recombinant protein contained A-domain which has the properties of binding to a heparin- related ligand on HepG2 liver cells. This current observation on recombinant and refolded PvTRAP firstly could be attributed to the possibility of the protein undergoing post-translational modifications in Escherichia coli, even though bacterial expression systems often fail to post-translationally modify a eukaryotic protein properly. Secondly, the role of A-domain earlier reported in Plasmodium TRAP which is present in our PvTRAP construct shares certain binding properties attributed to the region II plus like domain of TRAP in all Plasmodium species. As shown in our study, these properties could contribute to the observed binding of PvTRAP to heparin sulfate on hepatocytes. This observation clearly shows the importance of PvTRAP in the pre-erythrocytic invasion of liver cells by Plasmodium vivax.
The level of binding of the protein to HepG2 liver cells recorded in reduced and alkylated PvTRAP was comparatively less as compared to normal purified and refolded PvTRAP. The refolded PvTRAP had the disulfide bond structurally conformed, whereas, the reduced and alkylated PvTRAP had lost the disulfide bond by treatment with DTT and Iodoacetamide. Hence, this biochemical analysis affected the protein under the reduced and alkylated conditions by changing the structural conformation as compared to normally refolded and purified PvTRAP. This could be the basis for poor binding of the reduced and alkylated PvTRAP to HepG2 liver cells and demonstration of the protein specificity to these cells as shown under different biochemical conditions, as well as the migration and resolution of the protein on SDS-PAGE as shown in Figure 3b and 4.
Our experimental report on PvTRAP supports previous studies on binding of TRAP to sulfatide and heparin sulfate on the immortalized hepatocyte line HepG2 (14, 15, 35, 36), as well as other reported work on TRAP (37, 38). These results and the data on circular dichroism analysis on PvTRAP clearly demonstrated that the purified recombinant PvTRAP generated in our present investigation was properly refolded with good structural conformation, biological and functional activities.
Further analysis on the biological activity of purified and refolded PvTRAP was carried out using an erythrocyte interaction assay (Fig 5a&b). Purified and refolded recombinant PvTRAP recognized and interacted with Duffy-positive (D+) human erythrocytes, while no recognition and interaction was observed with Duffy- negative (D-) erythrocytes (Fig 5b). Positive binding was observed with Rhesus monkey, Rabbit and Mouse erythrocytes (Fig 5a &b). The interaction of recombinant PvTRAP with only human Duffy-positive erythrocytes is reported firstly in the present study. This data corroborated earlier findings on various studies on Plasmodium knowlesi, Plasmodium vivax and Plasmodium cynomolgi (41-43) and studies on binding properties of native P. vivax Duffy Binding protein (PvDBP) and region II of TRAP protein (32, 39). Though, region II of PvDBP bound to Aotus monkey erythrocytes, but no binding was reported with Rhesus monkey erythrocytes (32).
The present observation in our study revealed the interaction and recognition of PvTRAP with Rhesus monkey erythrocytes. However, it is important to note that the role of TRAP as a blood stage malaria parasite potential vaccine candidate is still not clear, but Plasmodium TRAP is a transmembrane protein whose part of its ectodomain consists of an A-domain and a thrombospondin type 1 repeat (TSR). Earlier studies on TRAP indicated the presence of 55-60 residues in TSR, five conserved residues and metal ion –dependent adhesion site (MIDAS) motif in A-domain (15, 16-19). The TSR and A domain of TRAP specifically interact with host cell receptors during cell invasion and these interactions are used by the parasite to exert force and actively penetrate the cell.
These TRAP adhesive modules are the only parasite ligands involved in productive interactions with the cell surface during cell invasion (15), therefore, purified and refolded recombinant PvTRAP which we expressed and reported here consists of these modules and they could act as a dual ligand system which appears to be important and sufficient for interaction and recognition of erythrocytes as well as potential for cell invasion (20-24). The adhesive modules present in this current protein- PvTRAP, most especially metal ion-dependent adhesion site motif which was made up of five residues, could play a critical role in ligand-binding and erythrocyte adhesive properties of recombinant PvTRAP. Our present observation on erythrocytic binding properties of PvTRAP may seem controversial, but it has generated the focus on the need to carry out further studies on the role of PvTRAP gene in erythrocytic Plasmodium vivax cell invasion and immuno-pathogenicity in man.
In conclusion, we have been able to express, purify and characterize refolded recombinant PvTRAP. The biochemical and binding properties of the protein clearly demonstrated the potential of PvTRAP as a good target and strong vivax malaria vaccine candidate.
|
|
|
|
|
|
|
|
|
ACKNOWLEDGEMENT |
|---|
Jamiu A. Ogunbanwo was a Research Fellow at the Nigerian Institute of Medical Research-NIMR, Lagos Nigeria and recipient of a UNIDO/ICGEB, Italy Research Fellowship Training Award (F/NIG 98-02). He carried out the present independent study under the supervision of Prof. V.S. Chauhan at ICGEB, Component in New Delhi, India.
We are grateful to Rakesh Kumar Singh and other research staff at ICGEB, New Delhi for their co-operation. We thank Puran S. Sijwali, Andrew Lynn and Sanjay Singh for expert technical support.
|
|
REFERENCES
|
|---|
1a. Lim CS, Tazi L, Ayala FJ. Plasmodium vivax: Recent world expansion and genetic identity to Plasmodium simium. PNAS 2005; 102 (43), 15523-15528.
1b. Jalah R, Sarin R, Sud N, Alam MT, Parikh N, Das TK, Sharma YD. Identification, expression, localization and serological characterization of a tryptophan-rich antigen from the human malaria parasite Plasmodium vivax. Mol. Biochem. Parasitol. 2005; 142: 158-169.
1c. Yang C, Collins WE, Sullivan JS, Kaslow DC, Xiao L, Lal AA. Partial Protection against Plasmodium vivax Blood- stage Infection in Samiri Monkeys by Immunization with a Recombinant C-Terminal Fragment of Merozoite Surface Protein 1 in Block Copolymer Adjuvant. Infect Immun 1999; 342- 9.
2. Arnot DE, Barnwell JW, Tam JP, Nussenzweig V, Nussenzweig RS, Enea V. Circumsporozoite protein of Plasmodium vivax: gene cloning and characterization of the immunodominant epitope. Science 1985; 230: 815-18.
3. Fang X, Kaslow DC, Adams JH, Miller LH. Cloning of the Plasmodium vivax duffy receptor. Mol Biochem Parasitol 1991; 44: 125-32.
4. Lakshman Perera KLR, Handunnetti SM, Holm I, Longacre S, Mendis KN. Baculovirus merozoite surface protein 1 C-terminal recombinant antigens are highly protective in a natural primate model for human Plasmodium vivax malaria. Infect Immun 1998; 66: 1500- 06.
5. Snewin VA, Khouri E, Wattavidanage J, Perera L, Premawansa S, Mendis KN, David PH. A new polymorphic marker for PCR typing of Plasmodium vivax parasites. Mol Biochem Parasitol 1995; 71: 135-8.
6. Rogers WO, Malik A, Mallouk S, Nakamura K, Rogers MD, Szarfman A, Gordon DM, Nusselvi AK, Aikawa M, Hoffman SL. Characterization of Plasmodium falciparum sporozoites surface protein 2. Proc Natl Acad Sci USA 1992; 89: 9176-80.
7. Cowan G, Krishna S, Crisanti A, Robson K. Expression of thrombospondin- related anonymous protein in Plasmodium falciparum sporozoites. Lancet 1992; 339: 1412-3.
8. Robson KJH, Frevert U, Reckman I, Cowan G, Beier J, Scragg IG,Takehara K, Bishop DHL, Pradel G, Sinden R, Saccheo S, Muller HM, Crisanti A. Thrombospondin related adhesive protein (TRAP) of Plasmodium falciparum expression during sporozoites ontogeny and binding to human hepatocytes. EMBO J 1995; 14 (16): 3883-94.
9. Templeton TJ, Kaslow DC. Cloning and cross species comparison of the thrombospondin-related anonymous protein (TRAP) gene from Plasmodium knowlesi, Plasmodium vivax and Plasmodium gallinaceum. Mol Biochem Parasitol 1997; 84: 13-24.
10. Sijwali PS, Malhotra P, Puri SK, Chauhan VS. Cloning and sequence analysis of the thrombospondin- related adhesive protein (TRAP) gene of Plasmodium cynomolgi bastianelli. Mol. Biochem. Parasitol. 1997; 90: 371-6.
11. Jongwutiwes S, Putaporntip C, Kanbara H, Tanabe K. Variation in the thrombospondin-related adhesive protein (TRAP) gene of Plasmodium falciparum from Thai field isolates. Mol Biochem Parasitol 1998; 92: 349-53.
12. Yuda M, et al. Targeted disruption of the Plasmodium berghei CTRP gene reveals its essential role in malaria infection of the vector mosquito. J Exp Med 1999; 190: 1711-15.
13. Dessens JT et al. CTRP is essential for mosquito infection by malaria ookinetes. EMBO J 1999; 18: 6221-7.
14. Nussenzweig V, Menard R. Analysis of a malaria sporozoite protein family required for gliding motility and cell invasion. Trends in Microbiology 2000; 8 (3): 1-3.
15. Menard R. The journey of the malaria sporozoite through its hosts: two parasite proteins lead the way. Microbes and Infection 2000; 2: 633-42.
16. Lee JO, Rieu P, Arnaout MA, Liddington R. Crystal structure of the A domain from the a subunit of integrin CR3 ( CD11b/CD18). Cell 1995; 80: 631-8.
17. Lee JO, Bankston LA, Arnaout MA, Liddington R. Two conformations of the integrin A-domain (I-domain): a pathway for activation? Structure 1995; 3: 1333-40.
18. Qu A, Leahy DJ. Crystal structure of the I-domain from the CD11a/CD18 (LFA-1, αLβ2) integrin. Proc Natl Acad Sci USA 1995; 92: 10277-81.
19. Qu A, Leahy DJ. The role of the divalent cation in the structure of the I domain from the CD11a/CD18 integrin. Structure 1996; 4: 931- 42.
20. Michishita M, Videm V, Arnaout MA. A novel divalent cation-binding site in the A domain of the β2 integrin CR3 (CD11b/CD18) is essential for ligand binding. Cell 1993; 72: 857- 66.
21. Kern A, Briesewitz R, Bank I, Marcantonio EE. The role of the I domain in ligand binding of the human integrin α1β1. J Biol Chem 1994; 269: 22811-6.
22. kamata T, Takada Y. Direct binding of collagen to the I domain of integrin α2β1 (VLA-2,CD49b/CD29) in a divalent cation-independent manner. J Biol Chem 1994; 269: 26006-10.
23. Ueda T, Rieu P, Brayer J, Arnaout MA. Identification of the complement iC3b binding site in the β2 integrin CR3 (CD11b/CD18). Proc Natl Acad Sci USA 1994; 91: 10680-4.
24. Kamata T, Wright R, Takada Y. Critical threonine and aspartic acid residues within the 1 domains of β2 integrins for interactions with intercellular adhesion molecule 1 (ICAM-1) and C3bi. J Biol Chem 1995; 270: 12531-5.
25. Scarselli E, Tolle R, Koita O, Diallo M, Muller HM, Fruh K, Doumbo O, Crisanti A, Bujard H. Analysis of the human antibody response to thrombospondin- related anonymous protein of P. falciparum. InfectImmun 1993; 61: 3490-5.
26. Sharma P, Bharadwaj A, Bhasin VK, Sailaja VN, Chauhan VS. Antibodies to a conserved-motif peptide sequence of the P. falciparum thrombospondin- related anonymous protein and circumsporozoite protein recognize a 78-KDa protein in the asexual blood stages of the parasite and inhibit merozoite invasion in vitro. Infect Immun 1996; 64: 2172-9.
27. Tung Pradubkul S, Panyinm S. isolation of DNA and RNA. In: Panyim S, Wilarat P, Yuthavong Y- editors. Application of Genetic Engineering to Research on Tropical Disease pathogens with Special Reference to Plasmodia. Geneva: WHO. 1985; 51-56.
28. Simons PC, VanderJagt DL. Purification of glutathione S-transferases from human liver by glutathione-affinity chromatography. Anal Biochem 1977; 82: 334-41.
29. Chang M, Bolton JL, Blond SY. Expression and purification of Hexahistidine- Tagged Human Glutathione S- Transferase P1 in Escherichia coli. Prot Exp Pur 1999; 17: 443-8.
30. Glansbeek HL, VanBeningen HM, Vitters EL, Van der Kraan PM, Van der Berg WB. Expression of Recombinant Human Soluble Type II Transforming Growth Factor-beta Receptor in Pichia pastoralis and Escherichia coli: Two Powerful Systems to Express a Potent Inhibitor of Transforming Growth Factor-beta. Prot Exp Pur 1998; 12: 201-207.
31. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 1976; 72: 248-54.
32. Dutta S, Daugherty JR, Ware LA, Lanar DE, Ockenhouse CF. Expression, purification and characterization of a functional region of the Plasmodium vivax Duffy binding protein. Mol Biochem Parasitol 2000; 109: 179-84.
33. Reed MB, Caruana SR, Batchelor AH, Thompson JK, Crabb BS, Cowman AF. Targeted disruption of an erythrocyte binding antigen in Plasmodium falciparum is associated with a switch toward a sialic acid – independent pathway of invasion. Proc. Natl Acad Sci 2000; (97) 13: 7509-14.
34. Rogers WO, Rogers MD, Hedstrom RC, Hoffman SL. Characterization of a gene encoding sporozoite surface protein 2, a protective Plasmodium yoelii sporozoite antigen. Mol Biochem Parasitol 1992; 53: 45-52.
35. McCormick CJ, Tuckwell DS, Crisanti A, Humphries MJ, Hollingdale MR. Identification of heparin as a ligand for the A-domain of Plasmodium falciparum thrombospondin- related adhesion protein. Mol Biochem Parasitol 1999; 100: 111-124.
36. Wengelnik K, Spaccapelo R, Naitza S, Robson KJH, Janse CJ, Bistoni F, Waters AP, Crisanti A. The A-domain and the thrombospondin-related motif of Plasmodium falciparum TRAP are implicated in the invasion process of mosquito salivary glands. EMBO J 1999; (19) 18: 5195-5204.
37. Wengelnik K, Spaccapelo R, Naitza S, Crisanti A. Analysis of a malaria sporozoite protein family required for gliding motility and cell invasion: Response. Trends in Microbiology 2000; (8) 3: 1-2.
38. Matuschewski K, Nunes AC, Nussenzweig V, Menard V. Plasmodium sporozoite invasion into insect and mammalian cells is directed by the same dual binding system. EMBO J 2002; (21) 7: 1597-1606.
39. Wertheimer SP, Barnwell JW. Plasmodium vivax interaction with the human Duffy blood group glycoprotein: identification of a parasite receptor- like protein. Exp Parasitol 1989; 69: 340-50.
40. Vokel D, Blankenfeldt W, Schomburg D. Large- scale production, purification and refolding of the full-length prion protein from Syrian golden hamster in Escherichia S- transferase-fusion system. Eur J Biochem 1998; 251: 462-71.
41. Miller LH, Mason SJ, Dvorak JA, McGinnis MH, Rothman IK. Erythrocyte receptors for (Plasmodium knowlesi ) malaria: Duffy blood group determinants. Science 1975; 189: 561.
42. Miller LH, Mason SJ, Clyde DF, McGinnis MH. The resistance factor to Plasmodium vivax in blacks: Duffy blood group genotype FyFy. New Engl J Med 1976; 295: 302.
43. Okenu DMN, Malhotra P, Lalitha PV, Chitnis CE, Chauhan VS. Cloning and sequence analysis of a gene encoding an erythrocyte binding protein from Plasmodium cynomolgi. Mol Biochem Parasitol 1997; 89: 301-6.
44. Espinosa AM, Sierra AY, Barrero CA, Cepeda LA, Cantor EM, Lombo TB, Guzman F, Avila SJ, Patarroyo MA. Expression, polymorphism analysis, reticulocyte binding and serological reactivity of two Plasmodium vivax MSP-1 protein recombinant fragments. Vaccine 2003; 21: 1033-1043.
45. Patarroyo MA, Perez-Leal O, Lopez Y, Cortes J, Rojas-Caraballo J, Gomez A, Moncada C, Rosas J, Patarroyo ME. Identification and characterization of the Plasmodium vivax rhoptry-associated protein 2. Biochem and Biophys Res Comm. 2005; 337: 853-859
46. Gaffar FR, Yatsuda AP, Franssen FFJ, deVries E. A Babesia bovis merozoite protein with a domain architecture highly similar to the thronbospondin- related anonymous protein (TRAP) present in Plasmodium sporozoites. Mol. Biochem. Parasitol. 2004; 136: 25-34.
47. Altstein AD, Gitelman AK, Smirnov YA, Piskareva LM, Zakharova LG, Pashvykina GV, Shmarov MM, Zhirnov OP, Varich NP, Ilyinski PO, Shneider AM. Immunization with influenza A NP-expressing vaccinia virus recombinant protects mice against experimental infection with human and avian influenza viruses. Arch. Virol. 2006; 151:921-931.