| REVIEW ARTICLE |
|
|
1 Department of Physiology, School of Basic Medicine, Qingdao University, Qingdao, China;
2 Institute of Brain Science and Disorders, Qingdao University, Qingdao, China
Corresponding Author: Jie Wu, Professor, Institute of Brain Science and Disorders, Qingdao University, Room 509, Boya building, 308 Ningxia Rroad, Qingdao, 266071, China. E-mail: jiewu2@qdu.edu.cn.
Note: MengZhen Wang and Sha Zhao contributed equally.
| |
ABSTRACT |
| INTRODUCTION | |
|
|
INTRACELLULAR SECOND MESSENGER |
|
|
α7-nAChR AND INTRACELLULAR SECOND MESSENGER |
|
|
CONCLUSION |
|
|
AUTHOR CONTRIBUTIONS |
|
|
FUNDING |
|
|
CONFLICT OF INTERESTS |
|
|
ABBREVIATIONS |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Nicotinic acetylcholine receptors (nAChRs) are the members of the cys-loop ligandgated ion channel superfamily. They are cation-selective membrane proteins with a homopentameric or heteropentameric structure. The α7 subtype of nAChR (α7-nAChR) belongs to the classical homopentameric structure with the characteristics of relatively low affinity to agonists and high permeability to Ca2+. α7-nAChR plays an important role in modulating learning and memory, cognition, anti-inflammatory, growth and development, and other processes. There are many possible mechanisms underlying α7-nAChR-mediated modulations, including opening the high Ca2+ permeable channels, regulating presynaptic neurotransmitter release, and altering intracellular second messenger-mediated signaling pathways. Here, we summarize the signal pathways associated with α7-nAChR activation involving nucleotides, ions, lipids, and gas molecules. This article provides insights into α7-nAChR-mediated modulations through intracellular signaling systems and helps to understand the α7-nAChR-associated pathogenesis and therapeutics of a variety of diseases and disorders.
KEY WORDS: α7-nAChR; cAMP; Ca2+; Intracellular second messenger; IP3; NO; Signaling pathways|
|
INTRODUCTION |
|---|
Acetylcholine receptors
There are two main types of acetylcholine receptors (AChRs): muscarinic acetylcholine receptors (mAChRs) and nicotinic acetylcholine receptors (nAChRs) (1). Both subtypes are activated by the endogenous neurotransmitter acetylcholine (ACh), and they are expressed in neurons and non-neuronal cells, participating in various physiological functions (2, 3) . mAChRs are typical G protein-coupled receptors, divided into five subtypes: M1-M5 (4). nAChRs are ligand-gated ion channels and the activation of nAChRs results in rapid cellular responses. Therefore, the application of nAChR agonists usually does not produce or produce only a barely detectable response in various tissues and organs (5).
nAChR is a member of the cys-loop ligand-gated ion channel superfamily (1, 6) . They are cation-selective membrane proteins with a homopentameric or heteropentameric structure (7). The nAChRs are composed of different combinations of α (α2-α10) and β (β2-β4) subunits or α [7-9] subunit alone (8). The structure of nAChRs is consists of an extracellular domain, a transmembrane domain, and an intracellular domain. To date, more than 13 different neuronal nAChR subtypes have been discovered, including α1-α10, and β2-4 (1), each of which is closely related to a series of pathophysiological functions involving Parkinson's disease (PD), Alzheimer's disease (AD), and epilepsy, etc. (9-11) . In addition, α7-nAChR is associated with schizophrenia and AD (12) .
In Goodman & Gilman's "The Pharmacological Basis of Therapeutics" (12th Edition, 2011), nAChR is classified into muscle type (Nm), peripheral neuron type (Nn), and central neuron type (CNS) based on the distribution, subunit composition, and selective antagonists. CNS nAChRs are expressed in neurons and glial cells in numerous brain regions. In its classification, the CNS nAChRs are further divided into two subtypes according to whether they are sensitive to α-bungarotoxin: (α4)2 (β2)3(insensitive) and (α7)5(sensitive). In all types of nAChRs, the opening of ion channels induced by agonists (such as ACh or nicotine) binding causes Na+ and Ca2+ influx. This depolarizes the cells and opens various function switches (1). Detecting postsynaptic nicotine responses of neurons in the central nervous system is relatively complicated, as most neuronal nAChRs rapidly desensitize when exposed to nicotine agonists (1, 13) .
α7 nicotinic acetylcholine receptors
The neuronal type α7-nAChR is a homopentamer, meaning that it consists of five identical subunits. α7-nAChRs are highly expressed in the central nervous system (cortex, striatum, and hippocampus) in the human brain (14) . They play an important role in advanced neural functions (such as cognitive function, learning, and memory processes) and cholinergic anti-inflammatory pathways (15-17) . Emerging evidence suggests that the altered α7-nAChR subtype is related to the pathogenesis of AD (18) .
Natural α7-nAChRs are expressed on both presynaptic and postsynaptic membranes, and they act as the regulators of circuit activity in the brain (8, 19) . Although α7-nAChRs are desensitized in the sub-millisecond range, its relatively high Ca2+ permeability exhibits physiological significance (20) . In fact, selective activation of α7-nAChRs indicates that they improve the learning and memory, and anxiety conditions (21, 22) . On the other hand, deficits of α7-nAChR function are associated with various diseases including AD, schizophrenia and pain (23, 24) . However, accumulating lines of evidence suggest that up-regulation of α7-nAChRs may produce neurotoxic effects and damage neurons (25-27) . These lines of evidence suggest that the α7-nAChR-mediated modulations may involve complicated mechanisms and signal pathways.
|
|
INTRACELLULAR SECOND MESSENGER |
|---|
The second messengers are a series of small molecules and ions that respond to the first messenger outside the cell and transmit the signal received by the cell surface receptor to the effector protein (28, 29) . Its synthesis and degradation are controlled by a variety of enzymes in mammalian cells (28). Since the discovery that cyclic adenosine monophosphate (cAMP) (the first found second messenger) plays a role in the signal transduction process (30), more small molecules and ions have been found to also play a similar role, and have different nature, allowing them to carry information within the membrane (hydrophobic molecules, such as lipids and lipid derivatives), in the cytoplasm (polar molecules, such as nucleotides and ions), or between the two (such as gases and free radicals) transfer (29, 31) . Second messengers not only respond to extracellular stimuli, but also respond to intracellular stimuli. They usually are available in resting cells at lower concentrations and can be produced or released quickly when cells are stimulated (29). The concentration level of second messengers is precisely regulated by diverse mechanisms. In the process of information transmission, enzymatic reactions or the opening of ion channels ensure that they are highly amplified to improve the accuracy of cell signal transduction (32) . There are only a few known second messengers, but they can transmit a variety of extracellular information, cause-specific intracellular reactions, and thereby regulate different physiological and biochemical processes (31).
|
|
a7-nAChR AND INTRACELLULAR SECOND MESSENGER |
|---|
α7-nAChR and cAMP
Adenylate cyclase (AC) convert adenosine triphosphate (ATP) into cAMP and pyrophosphate. AC can be divided into three families: calcium ion and calmodulin activated type (AC1, AC3, and AC8), Ca2+ inhibited type (AC5, AC6, and AC9) and Ca2+ insensitive type (AC2, AC4, and AC7) (33) .
It is well known that the activation of AC can increase the release of glutamate from hippocampal neurons (34) . For a long time, cAMP has been recognized as a gated molecule that controls the expression of short-term plasticity and its transformation into long-term plasticity (35, 36) . Cheng et al. (2011) investigated the relationship between cAMP and α7-nAChR-mediated signal transduction mechanism in cultured mouse hippocampal neurons and found that activation of α7-nAChRs resulted in an increased intracellular cAMP level, which depended on the increase of cytoplasmic calcium and calcium-stimulated AC1 activations (37) . It has been reported that the use of selective AC1 inhibitors or siRNA to inhibit AC1 can prevent α7-nAChR-mediated cAMP elevation, which indicates that calcium-dependent AC1 can convert short-term calcium elevation into cAMP signal when α7-nAChR is activated, and then lead to a series of cascading reactions (38) .
The activation of α7-nAChRs induced the prolongation and enhancement of glutamatergic synaptic transmission in a PKA-dependent manner (39) . In the presence of α7-nAChR positive allosteric modulator PNU-120596, treatment with α7-nAChR selective agonist choline can increase intracellular cAMP levels. This increase in cAMP induced by choline was eliminated by the α7-nAChR antagonist methyllycaconitine (MLA) and the calcium chelator BAPTA, indicating that the increase of cAMP depends on the activation of α7-nAChRs and the subsequent increase in intracellular calcium. Both the selective AC1 inhibitor CB-6673567 and siRNA-mediated AC1 deletion prevented the increase in cAMP brought about by choline, indicating that the effect of choline requires calcium-dependent AC1 (37, 38) .
In addition, in the hippocampus, calcium-stimulated presynaptic terminal AC-dependent PKA activation is mainly driven by AC1 rather than AC8 (40) . There is evidence that the activity of PKA is necessary for α7-nAChR-mediated enhancement of evoked excitatory post-synaptic current (eEPSC) amplitude. Previous studies have shown that PKA can regulate synaptic plasticity of the entire hippocampus, especially at the ends of mossy fibers (41-45) . Besides, inhibition of PKA activity can block the α7-nAChR-mediated effect on hippocampal synaptic plasticity (46) . It can be seen that under the stimulation of calcium and calmodulin, AC1 is a suitable candidate for coupling α7-nAChR with PKA (37, 39) .
To sum up, α7-nAChRs use the AC1-cAMP-PKA signal transduction pathway to phosphorylate synaptic proteins, and the synaptic proteins act as downstream effectors that regulate neurotransmitter release, thereby mediating its regulation of synaptic transmission and positive effects on cognition (37) .
α7-nAChR and Ca2+
It is well known that Ca2+ is an important signal transduction molecule in neuronal cells (47) . Many studies have also shown that α7-nAChR-mediated effects are mediated by the activation of intracellular Ca2+ signals, which reveals the physiological roles of receptor channels (48) .
Ca2+-AC1-cAMP-PKA/PKC. It has been reported that in cystic fibrosis disease (CF), loss or dysfunction of the transmembrane conductance regulator (CFTR) leads to an impaired airway mucus transport. The α7-nAChR is a key regulator of CFTR functional activity in airway epithelial cells of surface epithelium and submucosal glands, and the two are functionally coupled through Ca2+: Activation of α7-nAChR leads to Ca2+ influx, which leads to AC1 activation and cAMP production, which in turn initiates a series of PKA/PKC cascades, eventually leading to CFTR Cl- channel outflow of Cl-, and maintaining normal airway epithelium homeostasis (49, 50) .
Ca2+/CaM-eNOS-NO. There is evidence that a variety of neuromodulators are involved in the cardiovascular control of nucleus tractus solitarii (NTS), of which nicotine plays a key role in central cardiovascular control (51, 52) . eNOS is a constitutive and strictly Ca2+/calmodulin(CaM)-dependent enzyme. When the intracellular calcium concentration increases, calmodulin will replace caveolin, and act by stimulating phosphorylation of serine 1177 of eNOS (53) . Previous experiments have found that microinjection of nicotine into the NTS of Wistar-Kyoto rats (WKY rats) can cause a dose-dependent decrease in blood pressure (BP) and heart rate (HR), which indicates that nAChRs play an important role in the BP control of NTS. After pretreatment with nAChR antagonists or calmodulin-eNOS pathway blockers, this inhibitory effect of nicotine was weakened. In contrast, treatment with nNOS-specific inhibitors did not attenuate the nicotine-mediated effects. This indicates that eNOS may be one of the downstream targets of nAChRs, and regulate the BP in NTS of WKY rats by participating in the production of NO. And after nicotine was injected into NTS, it was found that calmodulin can bind to eNOS (47) . In short, the antihypertensive effect of nAChR regulation in NTS is achieved through the production of NO induced by eNOS, and the regulation of eNOS activity is achieved through the combination with Ca2+/CaM, that is, α7-nAChR-Ca2+/CaM-eNOS-NO pathway may play a role in the regulation of BP in NTS.
Ca2+–CaMKII–PKC. Acetylcholine concentrations in corneal epithelium are very high (54) , even exceeding the concentrations in nerve tissue (55) . ACh axis plays an important role in regulating and coordinating the different activities of corneal epithelial cells mediated re-epithelialization (56) . It was found that autocrine/paracrine ACh and cholinergic agonists could up-regulate the expression of E-cadherin by activating α7-nAChR, thus promoting corneal epithelial regeneration. Activation of α7-nAChRs can indirectly initiate the Ras–Raf–MEK–ERK cascade through the Ca2+–CaMKII–PKC signal pathway, or it can directly interact with Ras (57) .
Ca2+–CaM-CaMKII-CREB. AD is the most common type of neurodegenerative diseases, and there are multiple pathogenesis hypotheses, among which amyloid β protein(Aβ) deposition plays an important role in the pathogenesis of AD (18, 58-60) . Aβ deposition in the brain can lead to the imbalance of neuronal calcium homeostasis and decline in cognitive function (61) . nAChRs are involved in various important physiological processes, especially in learning, memory, and cognition, and α7-nAChRs is one of the subtypes of nAChR related to the pathogenesis of AD (18) . Also, studies have found that in different AD models, the use of α7-nAChRs-specific agonists can improve cognitive impairment (62) .
Studies have shown that cognitive decline in AD patients is closely related to impaired synaptic function (63). Synapses are the basic structure of connections between neurons, play a key role in information transmission, and are the foundation of learning and memory. The function of synapses is largely related to synapse-related proteins (18) . It has been found that the activation of α7-nAChRs by PNU-282987 (α7-nAChR specific agonist) can restore the expression levels of synapse-related proteins down-regulated due to Aβ deposition, including SYN, PSD95, SNAP25, DYN1, and AP180. When the α7-nAChR antagonist MLA was used to treat primary hippocampal neurons, the expression of the synapse-related protein decreased. As an intracellular second messenger, Ca2+ can get into the cell through α7-nAChRs and combine with CaM to form an active CaM complex. The CaM complex can bind to CaMKII, change the spatial conformation of CaMKII and phosphorylate it. Phosphorylated CaMKII enters the nucleus to catalyze the phosphorylation of CREB. CREB is located in the downstream of the α7-nAChR signal transduction pathway, and acts as a transcription factor to regulate the growth and development of neurons and the formation of long-term memory, thereby exerting neuroprotective effects (18, 64, 65) .
Changes in Ca2+ homeostasis in AD patients are related to synaptic dysfunction and neuronal decline (66) . PNU-282987 activates α7-nAChRs in both in vivo and in vitro models, thereby activating CaM-CaMKII-CREB signaling pathway to maintain intracellular Ca2+ steady state (18, 66) . This may provide a new understanding of the underlying pathogenesis of AD.
α7-nAChR and IP3
Nicotine activation of presynaptic nAChRs can promote neurotransmitter release and produce short-term and long-term enhancement effects on synaptic plasticity (67, 68) . Regulating the entry of Ca2+ at the presynaptic terminal is an important step to determine the degree of neurotransmitter release (69) . The nAChRs-mediated Ca2+ influx and subsequent Ca2+-dependent signal stimulation may be involved in the continuous phase of nicotine-induced synaptic transmission (67) . The activation of nAChRs can induce Ca2+ influx through direct penetration of receptor channels and activation of voltage-gated Ca2+ channels. Activation of α7-nAChR (high Ca2+ permeability) causes an increase in Ca2+ at the presynaptic terminal, which is sufficient to induce the release of calcium from the IP3-sensitive Ca2+ pool and maintain continuous changes in intracellular Ca2+ (69) . Activation of α7-nAChR can also activate phospholipase C (PLC) through a certain mechanism. PLC decomposes PIP2 to generate IP3, thereby mobilizing IP3 receptors to regulate calcium storage (70, 71) . CaMKII is activated by the activation of α7nAChR, and the activated CaMKII maintains the increase of presynaptic Ca2+ through a positive feedback mechanism. Activated CaMKII is related to presynaptic neurotransmitter vesicles and participates in the regulation of neurotransmitter release and synaptic transmission (72, 73) . The continuous changes of intracellular Ca2+ and the activation of CaMKII at the presynaptic terminal trigger the long-term enhancement of glutamate release and induce long-term promotion of glutamatergic synaptic transmission (69) .
After nicotine activates α7-nAChR, through the α7-nAChR-PLC-IP3-CaMKII cascade, or α7-nAChR directly interacts with Ca2+, both can trigger the release of presynaptic glutamate, which ultimately acts on the postsynaptic region, causing a series of physiological and biochemical reactions.
In addition, α7-nAChRs are also involved in signal transduction processes such as the activation of PLC and the release of intracellular Ca2+ in microglia, rather than acting as a traditional ion channel (70, 74) . This α7-nAChR signal transmission may be involved in the functional activation process of nicotine on microglia, exerting neuroprotective effects by inhibiting the inflammatory state and enhancing the protective function (70) .
α7-nAChR and NO
During the mutual regulation of nicotine and ethanol, nAChR acts as an intermediate target (75) . The cross-tolerance between nicotine and ethanol is dependent on the nAChR (76-78) . However, it is worth noting that α7-selective agonists can attenuate ethanol-induced ataxia, and this effect was partially block by α7-selective antagonists, which indicates that this process may be mediated by α7-nAChRs (78) . Pretreatment with RJR-2403 (α4β2nAChR selective agonist) or PNU-282987 (α7-nAChR selective agonist) can reverse the reduction of NOx (total nitrate + nitrite) caused by ethanol. The change of NOx level indicates that the NO-cGMP signaling pathway may be a mechanism of α4β2 and α7-nAChR subtypes mediated cross tolerance between nicotine and ethanol (75, 79) .
In short, α7-nAChR may participate in the interaction between nicotine and ethanol through the NO-cGMP signaling pathway, thereby reducing the ataxia caused by ethanol (78) .
|
|
CONCLUSION |
|---|
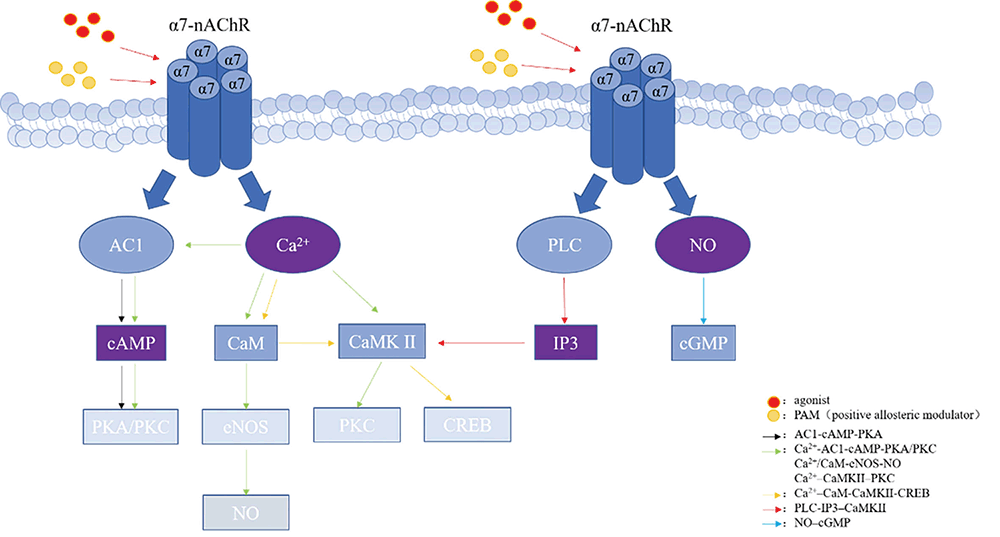
α7-nAChR is not only distributed in the central nervous system and peripheral nervous system, but also in the immune system. In recent years, there are more and more studies on the physiological or pathological roles of α7-nAChR. The wide distribution of α7-nAChR makes it a target for the treatment of neurodegenerative diseases and other diseases. Its agonists and positive allosteric modulators have also become the hot spots of drug research and development. We summarized that α7-nAChR can play a role through several intracellular second messengers and subsequent cascades (Table 1). We also found that the relationship between these signal cascades is complex (Figure 1). Further exploringe these signal networks, to understand the mechanisms and functions of α7-nAChR should be our focus.
|
|
|
|
AUTHOR CONTRIBUTIONS |
|---|
Jie Wu designed the outline study, analyzed information, and contributed to manuscript writing; Mengzhen Wang conducted the literature search and selection, then wrote the manuscript; Sha Zhao was responsible for summarizing tables and figures in articles; Yudan Zhang and Wei Liu contributed to study design. All of the authors have read and approved the final manuscript.
|
|
FUNDING |
|---|
This work was supported by the Key Area Research and Development Program of Guangdong Province (2018B030334001).
|
|
CONFLICT OF INTERESTS |
|---|
The authors declare that no conflicting interests exist.
|
|
ABBREVIATIONS |
|---|
| Aβ | amyloid β protein |
| AC | adenylate cyclase |
| Ach | acetylcholine |
| AChR | acetylcholine receptors |
| AD | alzheimer’s disease |
| ATP | adenosine triphosphate |
| BP | blood pressure |
| CaM | calmodulin |
| cAMP | cyclic adenosine monophosphate |
| CF | fibrosis disease |
| eEPSC | evoked excitatory post-synaptic current |
| HR | heart rate |
| mAChR | muscarinic acetylcholine receptors |
| MLA | methyllycaconitine |
| nAChR | nicotinic acetylcholine receptors |
| NTS | nucleus tractus solitarii |
| PD | parkinson’s disease |
| PLC | phospholipase C |
|
|
REFERENCES |
|---|