| REVIEW ARTICLE |
|
|
Crean Lutheran High School, 12500 Sand Canyon Ave, Irvine, CA 92618, USA
Corresponding Author: Jiaying Liu, Crean Lutheran High School, 12500 Sand Canyon Ave, Irvine, CA 92618, USA. E-mail: Yollie0424@gmail.com.
| |
ABSTRACT |
| INTRODUCTION | |
|
|
SONIC HEDGEHOG SIGNALING AND GLIOBLASTOMA CSCS |
|
|
POTENTIAL DRUGS TARGETING THE SONIC HEDGEHOG SIGNALING |
|
|
CONCLUSION |
|
|
ABBREVIATION |
|
|
ACKNOWLEDGEMENT |
|
|
CONFLICT OF INTEREST |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
Glioblastoma is an aggressive malignant brain tumor, and its five-year relative survival rate for patients is only 6.8 percent. In addition, almost all glioblastomas recur even after intensive treatments including surgery, radiation and chemotherapy, which are believed to be attributed to cancer stem cells (CSCs), a small population of cells existing in a tumor with ability to self-renewal and differentiation. Of multiple signaling pathways, sonic hedgehog signaling is one critical pathway involved in CSCs regulation. Therefore, targeting the sonic hedgehog signaling represents a novel therapy to treat glioblastomas, and significant efforts have been made in recent years to develop the sonic hedgehog signaling inhibitor drugs. This article specifically focuses on recent advances in the sonic hedgehog signaling inhibitor drug development by targeting multiple components of the signaling cascade including sonic hedgehog ligand, Smoothened (SMO) and GLI. Those inhibitors hold a great promise to destroy glioblastoma CSCs and prevent and treat tumor recurrence in the future.
KEY WORDS: sonic hedgehog; signaling; inhibitors; cancer stem cells; glioblastoma; SMO protein and GLI protein|
|
INTRODUCTION |
|---|
Glioblastoma, considered as the grade IV glioma based on World Health Organization report, is the most aggressive malignant brain tumor (1). The five-year relative survival rate of glioblastoma patients is only 6.8 percent, and the average length of survival for glioblastoma patients is estimated to be 12 to 18 months (2). Glioblastoma originates from astrocytes, the most abundant cell type in the central nervous system, and current treatment options include surgery, radiation, and chemotherapy as well as a target therapy antibody drug Bevacizumab (3, 4). However, none of those treatments can successfully extend patient lives beyond a few extra months, and glioblastoma recurrence and therapeutic resistance after the treatments are almost inevitable, which are believed to be attributed to CSCs (5). CSCs are a small population of cells existing in a tumor with ability to self-renewal and differentiation. CSCs were first identified from acute myeloid leukemia in 1994, and later have been reported in multiple tumor types including colon, breast, prostate, lung, melanoma, and glioblastoma (6, 7). CSCs play a significant role in tumor initiation and progression, and they can survive the traditional tumor treatments, accounting for therapeutic resistance and tumor relapse (8).
In 2006, CSCs was identified from glioblastoma with expression of CD133 marker, and the glioblastoma CSCs have been shown to survive chemotherapy drugs in cell-based assays and the murine models (5, 9, 10). Glioblastoma CSCs are difficult to target physically due to their presence in the perivascular space with very small population (11). While at the molecular level, activation of multiple signaling pathways has been reported to involve in glioblastoma CSCs’ properties including Wnt, Notch, and sonic Hedgehog signaling pathways (5). Thus, targeting these critical pathways at molecular level to kill glioblastoma CSCs represents a novel therapy to eradicate glioblastoma tumor, with a hope to overcome the therapeutic resistance and tumor reoccurrence issues faced by the traditional therapies. The important roles of the Wnt and Notch signaling pathways in glioblastoma CSCs can be found in the recent review papers (12, 13). This article specifically focuses on the recent advances in studying glioblastoma CSCs and drug development to target the sonic hedgehog signaling of CSCs for glioblastoma treatment.
|
|
SONIC HEDGEHOG SIGNALING AND GLIOBLASTOMA CSCS |
|---|
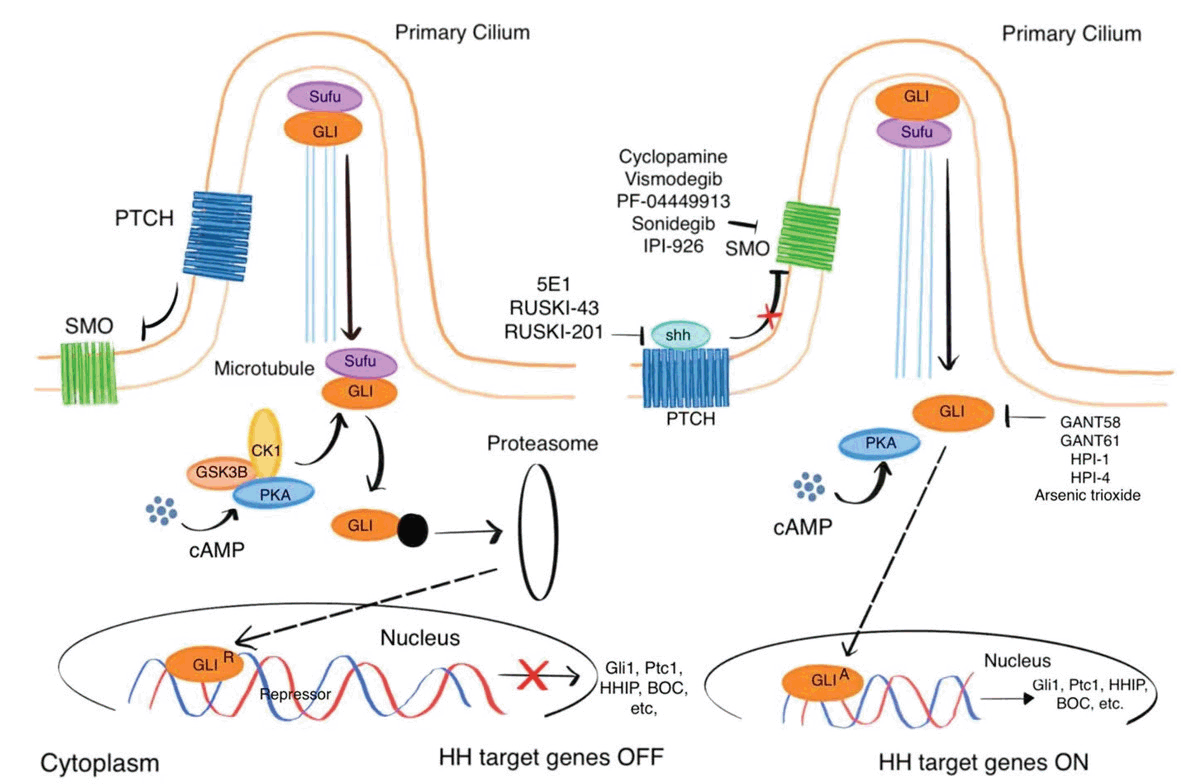
Sonic Hedgehog signaling is an essential pathway during vertebrate embryogenesis and they are responsible for organogenesis, regeneration and homeostasis (14, 15). In addition, abnormal activation of sonic hedgehog signaling is known to be implicated in many types of cancer (16). Collective evidence has demonstrated that sonic hedgehog signaling plays critical roles in CSCs’ stemness which accounts for tumor initiation, progression and recurrence as well as therapeutic resistance (17-19). In mammalian cells, the primary cilia (the thin and long organelles) protruding from the apical cellular surface, play an essential role in sonic Hedgehog signaling (20) . In the absence of the sonic hedgehog ligands, the transmembrane protein Patched (PTCH) is located on cilia to prevent the SMO protein accumulation to cilia (Figure 1). Under such condition, protein kinase A (PKA) can phosphorylate GLI transcription factor, and the phosphorylated GLI is then proteolyzed to repressor GLI (GLIR) which is subsequently translocated into nucleus to suppress the sonic hedgehog signaling target gene transcription (21) (Figure 1). However, in the presence of sonic HH ligands, the ligand binds to PTCH, which is moved out of the cilia and releases its inhibition to SMO. SMO is then activated and become enriched at primary cilia, leading to the activation of GLI which subsequently translocate into nucleus to induce target gene transcription (21, 22).
Abnormal activation of sonic Hedgehog signaling is associated with glioblastoma stem / progenitor cells. For instance, Barr et al found that the sonic hedgehog signaling is frequently upregulated in glioblastoma and inhibition of the sonic Hedgehog signaling reduces the cancer cell growth by depleting glioblastoma CSCs (18). The essential role of the sonic Hedgehog signaling in maintenance of glioblastoma CSCs is further confirmed by other groups (23, 24).
|
|
|
POTENTIAL DRUGS TARGETING THE SONIC HEDGEHOG SIGNALING |
|---|
In the past decades, significant research has been made to develop potential drugs to target the sonic Hedgehog signaling at distinct cascade levels.
Target the sonic Hedgehog signaling ligands
Sonic hedgehog ligands binding to PTCH receptor is the first step for the sonic hedgehog signaling activation, and therefore, targeting the sonic hedgehog ligands is a promising approach to block the hedgehog signaling. 5E1 is a monoclonal antibody which reacts with sonic hedgehog ligands, thus blocks the ligand binding to PTCH for the signaling inhibition (25). It has been demonstrated that 5E1 antibody effectively suppressed medulloblastoma and pancreatic tumors in respective mouse models (26, 27). In addition, RUSKI-43, a chemical compound, was developed by Petrova and colleagues to target the sonic hedgehog acyltransferase, an important enzyme for sonic hedgehog ligands processing before the ligands bind to the PCTH receptor (28). RUSKI-43 can lower growth of breast cancer cells and pancreatic tumor dramatically (29, 30). Nevertheless, RUSKI-43 was subsequently found to display cytotoxic activity. Rodgers et al has studied RUSKI-43 analogs and identified a RUSKI-201 with no off-target cytotoxicity and improved selectivity to sonic hedgehog acyltransferase (31). Whether 5E1 antibody and RUSKI-201 can effectively attenuate Glioblastoma CSCs by targeting the sonic hedgehog signaling remains unknown. Interestingly, chidamide (a histone deacetylase inhibitor) was found to inhibit the sonic hedgehog ligand expression by increasing miR-338-5p expression (32). As chidamide has been approved in China for the relapsed or refractory peripheral T-cell lymphoma and in Japan for the relapsed or refractory adult T-cell leukemia-lymphoma, further study of chidamide as a hedgehog signaling inhibitor for glioblastoma is warranted.
Targeting SMO protein
SMO is a crucial protein in the sonic hedgehog signaling pathway, and several SMO inhibitors have been developed in the past decades (33). In the famous Idaho cyclops lamb scenario, the Veratrum californicum was found to be responsible for the birth of cyclops lambs when the plant was consumed by pregnant sheep (34). Later, a steroidal alkaloid named cyclopamine (also known as 11-deoxojervine) was isolated from the Veratrum californicum, and cyclopamine was confirmed to block SMO protein activity for the inhibition of sonic hedgehog signaling. As the sonic hedgehog signaling is critical for embryonic development, inhibition of the sonic hedgehog signaling by cylcopamine leads to eye deformity (34). Subsequently, multiple studies have shown that cyclopamine can effectively reduce tumor growth by inhibiting the sonic hedgehog signaling (35). However, the application of cyclopamine for therapeutics is limited by its toxicity and poor solubility (35, 36). Subsequently, a cyclopamine derivative,IPI-926 ( also known as Saridegib) was developed, and IPI-926 targets SMO protein with much better metabolic stability, pharmacokinetics, and potency over cyclopamine (37). In addition, animal studies have demonstrated that IPI-926 effectively attenuated proliferation of several types of cancer such as medulloblastoma, chondrosarcoma, and pancreatic cancer (38-40). Vismodegib and Sonidegib are two SMO inhibitors that have been approved by FDA to treat basal cell carcinoma (41-43). Currently clinical trials of vismodegib, are ongoing for a variety of cancer types including acute myeloid leukemia and recurrent glioblastoma (35). Several studies have tested effects of Vismodegib and Sonidegib on CSCs in multiple types of cancer. For instance, Vismodegib was shown to impair the growth of CSCs in medulloblastoma and pancreatic cancer and colon cancer by inhibiting the sonic hedgehog signaling (44-46), while Sonidegib downregulates the expression of CSC markers and increases the sensitivity to paclitaxel in breast cancer (47). Recently, Huang et al have reported that inhibition of sonic hedgehog signaling by Sonidegib suppresses the proliferation of CD133+ glioma CSC likely through induction of autophagy (48).
Meanwhile, several other SMO inhibitors have been developed as well. For instance, Munchhof et al. identified PF-04449913 (glasdegib) which dramatically reduced the leukemia-initiation potential of acute myeloid leukemia cells in mice (49). Clinical study demonstrated that PF-04449913 is safe and well tolerant and efficacious for acute myeloid leukemia, myelofibrosis and advanced solid tumors (50-52). In 2019, PF-04449913 was approved by FDA to treat acute myeloid leukemia (53).
Targeting GLI protein
GLI is an essential transcription factor to mediate the sonic hedgehog signaling from cytoplasm to nucleus, and serves as a great target for the sonic hedgehog signaling inhibition. So far, three types of GLI inhibitors have been reported. GANT58 and GANT61 were identified in a cell-based screen and both of them block GLI1 expressions (54). GANT61 was shown to decrease the growth of prostate cancer and rhabdomyosarcoma in mouse tumor models (54, 55). Subsequently more studies have indicated that GANT61 dramatically downregulates the proliferation of colon cancer cells and ovarian cancer cells (56, 57).
Arsenic trioxide is another compound found to target GLI for the inhibition of the sonic hedgehog signaling (58). Beauchamp et al reported that arsenic trioxide exhibited cytotoxicity on Ewing sarcoma family of tumor and medulloblastoma cell lines in vitro and in vivo in the mouse models of Ewing sarcoma family of tumor and medulloblastoma (59). In addition, Nakamura et al studied the function of arsenic trioxide in osteosarcoma, and showed that arsenic trioxide can induce apoptotic cell death in osteosarcoma cells by inhibition of sonic hedge signaling (60). They further examined arsenic trioxide in mouse osteosarcoma model, showing that arsenic trioxide prevents the osteosarcoma growth in mice (60). As arsenic trioxide is an FDA-approved drug for leukemia treatment, it could become a new therapy for osteosarcoma. Moreover, a new class of sonic hedge pathway inhibitors termed HPI-1, HPI-2, HPI-3, and HPI-4 were identified by Hyman and colleagues in a high-throughput screening, which target GLI to effectively inhibit the sonic hedge signaling (61). Furthermore, Aspirin has been shown to sensitize malignant glioma cells to temozolomide chemotherapy by preventing the nuclear translocation of GLI1 to block the sonic hedgehog signaling. Therefore, combination of aspirin with temozolomide could be a promising strategy to treat malignant glioblastoma (62).
In summary, significant efforts have been made to develop the sonic hedgehog signaling inhibitors by targeting the signaling ligand, SMO and GLI, and three of them have been approved by FDA. They could be very valuable to destroy. glioblastoma CSCs.
|
|
CONCLUSION |
|---|
Glioblastoma is the most prevalent and lethal brain tumor, and maximal surgical resection followed by concurrent radiation or chemotherapy is the primary treatment option. However, it is almost inevitable that glioblastoma relapse and therapeutic resistance will occur after the treatment primarily owing to a small population of glioblastoma CSCs (5). Sonic hedgehog signaling of one of key pathways in regulation of self-renewal and differentiation of glioblastoma CSCs. Therefore, targeting the sonic hedgehog signaling is believed as an effective approach to destroy glioblastoma CSCs. Most of the recent developed drugs or inhibitors block the sonic hedgehog pathway by targeting the three key components, sonic hedgehog ligand, SMO and GLI.
To date, three drugs (Vismodegib, Sonidegib and Glasdegib) that inhibit the sonic hedgehog signaling by targeting SMO have been approved by FDA, and many other potential drugs to inhibit the sonic hedgehog signaling are under development. They represent a promising therapeutic approach to destroy CSCs in glioblastoma in the future. Nevertheless, it is noteworthy that the sonic hedgehog signaling is not the only pathway involved in regulation of the glioblastoma CSCs. Therefore, targeting multiple pathways essential to CSCs simultaneously by combined therapies may yield the best results.
|
|
ABBREVIATION |
|---|
CSCs |
Cancer Stem Cells |
SMO |
Smoothened protein |
PTCH |
Patched Protein |
PKA |
Protein Kinase A |
GLI |
Glioma-associated Oncogene |
HH acyltransferase |
Hedgehog Acyltransferase |
FDA |
U.S. Food and Drug Administration |
PF-04449913 |
Glasdegib |
5E1 |
Monoclonal Antibody |
|
|
ACKNOWLEDGEMENT |
|---|
I greatly appreciate the help of Dr. Jay Hao during the conception and process of this manuscript.
|
|
CONFLICT OF INTEREST |
|---|
The authors declare that no conflicting interests exist.
|
|
REFERENCES |
|---|