| Original Article |
|
|
1 Departamento de Microbiologia Geral, Instituto de Microbiologia Prof Paulo de Góes (IMPPG), Centro de Ciências da Saúde (CCS), Bloco I, Universidade Federal do Rio de Janeiro (UFRJ), Ilha do Fundão, 21941-590, Rio de Janeiro, RJ, Brazil.
2 Laboratório de Ultraestrutura Celular, Departamento de Ultraestrutura e Biologia Celular, Instituto Oswaldo Cruz, FIOCRUZ, Avenida Brasil 4365, 21045-900, Rio de Janeiro, RJ, Brazil.
Corresponding Author: Alane B. Vermelho, Departamento de Microbiologia Geral, Instituto de Microbiologia Prof Paulo de Góes (IMPPG), Centro de Ciências da Saúde (CCS), Bloco I, Universidade Federal do Rio de Janeiro (UFRJ), Avenida Carlos Chagas Filho 373, Ilha do Fundão, 21941-902, Rio de Janeiro, RJ, Brazil. Tel: +55 (21) 2562 6740; Fax: +55 (21) 2560 8344; E-mail: abvermelho@micro.ufrj.br.
Running title:Peptidases in non-infected and T. cruzi-infected hepatocytes
| |
ABSTRACT |
| INTRODUCTION | |
|
|
MATERIALS AND METHODS |
|
|
RESULTS AND DISCUSSION |
|
|
ACKNOWLEDGEMENTS |
|
|
REFERENCES |
|
|
ABSTRACT
|
|---|
|
|
INTRODUCTION |
|---|
Trypanosoma cruzi, the etiological agent of Chagas’ disease, is an intracellular protozoan parasite able to invade and multiply inside a wide variety of mammalian tissues and cells. T. cruzi employs multiple molecular strategies to invade a broad range of nonphagocytic cells (1), one of them is the expression of peptidases that play relevant roles in different phases of the parasite-host relationship. In this context, several proteolytic enzymes have been extensively studied in T. cruzi, including cysteine-, serine-, threonine- and metallo-type peptidase classes (2, 3). The most abundant and the best-characterized among these enzymes is cruzipain, a cysteine peptidase expressed as a complex mixture of isoforms by all the developmental stages of the parasite, including some membrane-bound isoforms. The enzyme is an immunodominant antigen in human chronic Chagas’ disease and seems to be crucial for parasite growth, development and tissue ⁄ host cell penetration. For these reasons, cruzipain has been proposed as a therapeutic target for treatment of Chagas’ disease (reviewed in 2).
Along the intracellular development of T. cruzi, the expression of extracellular matrix components was demonstrated in the evolution of the fibrosis process in a murine model (4). For this reason, the role of peptidases that are capable of degrading the components of the extracellular matrix, such as matrix metallopeptidases (MMPs), during the T. cruzi invasion process as well as in the regulation of pathogenic conditions seems to be of extreme significance. In this context, our laboratory detected the active (85 kDa) and latent (100 kDa) form of MMP-9, using a rabbit anti-mouse MMP-9 antibody in western blotting and immunocytochemistry analyses in T. cruzi-infected hepatocyte cell cultures. Conversely, only the latent form of MMP-9 was detected in non-infected hepatocytes (5). In the present work, we have evaluated the ability of the cellular peptidases, detected in the non-infected and in the T. cruzi-infected primary cultures of mouse embryo hepatocytes, in degrading different protein substrates. In addition, we looked for the detection of cruzipain molecules during its in vitro interaction.
|
|
MATERIALS AND METHODS |
|---|
Hepatocyte culture
Hepatocytes were isolated from mouse embryo livers according to the methodology described previously, with some adaptations (6, 7). Briefly, ten mouse embryo livers were asseptically removed and washed with Hank’s balanced salt solution Ca+2 and Mg+2 free (HBSS-CMF). Longitudinal cuts were made in each lobe for enzyme access and the livers were then incubated at 37 ºC for 20 min with 0.05% collagenase (Type II Wortington) in approximately 50 ml HBSS-CMF. The organs were dispersed by pipetting, and liver cells were collected by centrifugation at 200 g for 3 min. Viable cells were purified by sedimentation at room temperature for 10 min with MEM/199 medium containing 10% fetal calf serum (FCS). Cells yield was 2 × 107 ml (total 10 ml) and viability greater than 90%, as assessed by the trypan blue exclusion methodology. The cells were seeded on tissue culture flasks. Four hours later fresh defined medium [MEM/199 supplemented with 5 mM CaCl2, 10 mg/ml insulin, transferrin and selenium (ITS), 1 ng/ml glucagon, 50 ng/ml epidermal growth factor (EGF), 3.5 × 10-6 M hydrocortisone and 1 ng/ml bovine serum albumin (BSA)] was added and replaced daily. The cultures were incubated at 37 ºC in a humidified atmosphere containing 5% CO2.
Parasite
Bloodstream trypomastigotes of T. cruzi Y strain were obtained from seven days-infected mice in the peak of parasitemia. The blood was collected with 3.8% sodium citrate as an anticoagulant agent, and centrifuged at 150 g for 10 min. The tube was then incubated at 37 oC for 30 min, so that trypomastigotes could move from the pellet to the supernatant. The pellet was discarded and the supernatant, which contained the parasites, was collected, centrifuged at 900 g for 10 min and washed with cold PBS (150 mM NaCl; 20 mM phosphate buffer, pH 7.2) (8). Cell number was estimated by counting the parasites in a Neubauer chamber. Amastigote forms were obtained from the supernatant of Vero cell cultures infected with bloodstream trypomastigotes as previously described by Souto-Padrón et al. (9).
Parasite-hepatocyte interaction
The hepatocyte primary cultures were infected 48 h after plating with 5 × 106 parasites per flask. The cultures were incubated during 48 h at 37 ºC in a humidified atmosphere containing 5% CO2. A four-days non-infected hepatocyte culture was used as a control system (5, 6).
Hepatocyte extracts
The cells of both systems were scrapped from flasks, centrifuged at 2000 gfor 20 min at 4 ºC, washed three times with cold PBS, and then lysed at room temperature by the addition of 100 ml SDS-PAGE sample buffer (125 mM Tris, pH 6.8; 4% SDS; 20% glycerol; 0.002% bromophenol blue). Alternatively, the non-infected and the T. cruzi-infected hepatocyte culture supernatants were collected, and passed over a 0.22-mm filtration unit (Millipore). The samples were concentrated by dialysis (cut-off 9000 Da) against polyethyleneglycol 4000 overnight at 4 ºC. Protein concentration was determined by the method described by Lowry et al. (10), using BSA as standard. Finally, the SDS-PAGE sample buffer was added in a 7:3 (v/v) proportion.
Substrate-gel electrophoresis
The proteolytic activities in the culture supernatants (50 mg protein) and in the cellular extracts (1 × 106 hepatocytes) were evaluated by 10% SDS-PAGE containing 0.1% gelatin, casein or hemoglobin as substrates incorporated into the gel (11). After electrophoresis at a constant voltage of 160 V at 4 ºC, the gels were soaked for 1 h at 25 ºC in 2.5 % Triton X-100. Gels were then incubated for 24 h at 37 ºC in 50 mM sodium phosphate buffer, pH 5.5, supplemented with 2 mM dithiothreitol (DTT) in the absence or in the presence of 10 mM E-64, and in 50 mM glycine-NaOH buffer, pH 10, in the absence or in the presence of 10 mM 1,10-phenanthroline. The gels were stained for 1 h with 0.2% Coomassie brilliant blue R-250 in methanol-acetic acid-water (50:10:40) and destained in the same solvent (20). Molecular masses of the proteinases were calculated by comparison of the mobility of GIBCO BRL SDS-PAGE standards (Grand Island, NY, USA). The gels were dried, scanned and densitometric analysis was performed with the use of the Kodak Digital Science EDAS 120 software as described by Soares et al. (12).
Western-blotting
Cellular protein extracts (equivalent to 100 μg) from non-infected and T. cruzi-infected hepatocytes as well as amastigote forms of T. cruzi Y strain (50 μg), the intracellular replicative stage observed during mammalian infection, were electrophoretically transferred (100 V/300 mA for 2 h at 4 oC) to a nitrocellulose membrane. The membrane was blocked in 10% nonfat dried milk in a blocking solution (150 mM NaCl; 10 mM Tris, pH 7.5; 0.05% Tween 20) for 1 h at room temperature under constant agitation and washed three times (10 min per wash) in the same solution. The membrane was incubated with anti-cruzipain polyclonal antibody kindly provided by Dr. Juan-Jose Cazzulo, Instituto de Investigaciones Biotecnologicas, Universidad Nacional de General San Martin, Buenos Aires, Argentina) at 1:100 dilution for 2 h and then washed as described above. Finally, the blot was incubated with the secondary antibody for 1 h, followed by immunodetection by chemiluminescence (ECL reagent) (12).
|
|
RESULTS AND DISCUSSION |
|---|
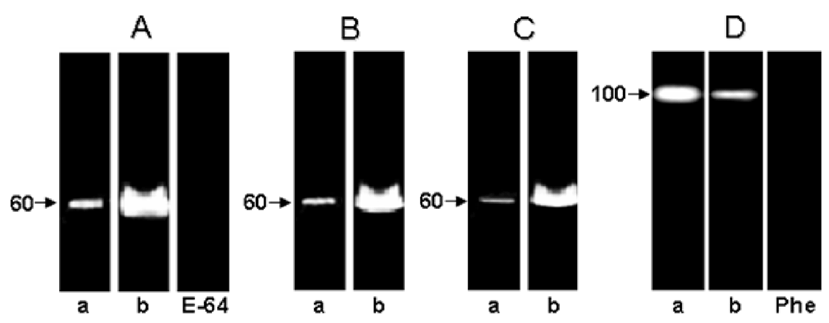
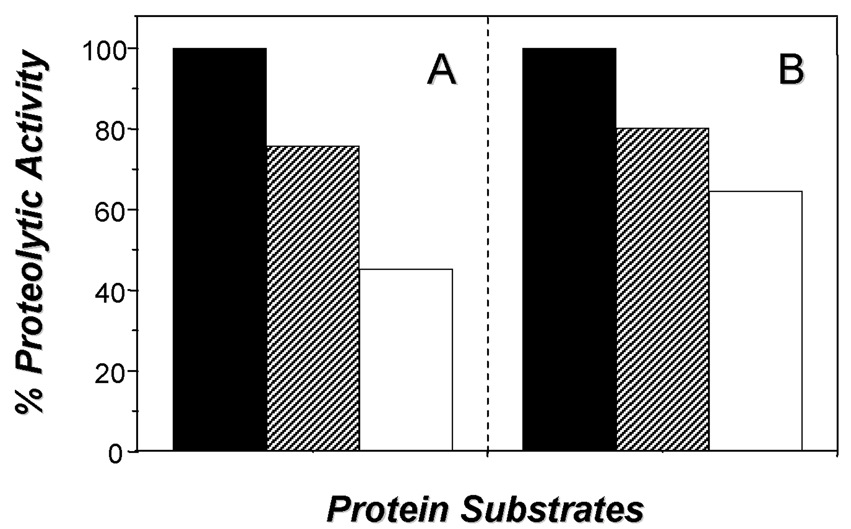
Trypanosoma cruzi has a complex life cycle comprising different developmental stages, which permits its adaptation to very diverse host environments. The tropism of T. cruzi for different tissues, including liver, has been reported (5, 7, 13). A recent study showed invasion and development through the entire T. cruzi cell cycle in an in vitro system of primary cultures of mouse embryo hepatocytes (7), affecting the hepatocyte peptidase production (5). In the present report, the hydrolytic ability of the peptidases present in these culture systems, infected or not with T. cruzi, was evaluated by SDS-PAGE containing different proteinaceous substrates. The zymogram gels showed a cell-associated peptidase migrating at 60 kDa in gelatin-SDS-PAGE. This proteolytic activity was detected in the control and infected hepatocyte cellular extracts only at acidic pH value (5.5) and in the presence of a reducing agent (DTT) (Fig. 1A, lanes a,b), being its hydrolytic activity completely abolished by 10 μM E-64, a potent cysteine peptidase inhibitor (Fig. 1, lane E-64). The 60 kDa cysteine peptidase presented the broadest specificity, since it was also able to degrade casein (Fig. 1B) and hemoglobin (Fig. 1C), although significantly less (Figs. 1 and 2). Table 1 summarizes the densitometric measurements of the proteolytic activities expressed in both hepatocyte systems using gelatin as substrate. Additionally, we observed that the activity of the 60 kDa cysteine peptidase was significantly enhanced (approximately 232%) in the T. cruzi-infected system (compare lanes a and b in the Fig. 1, and Table 1).
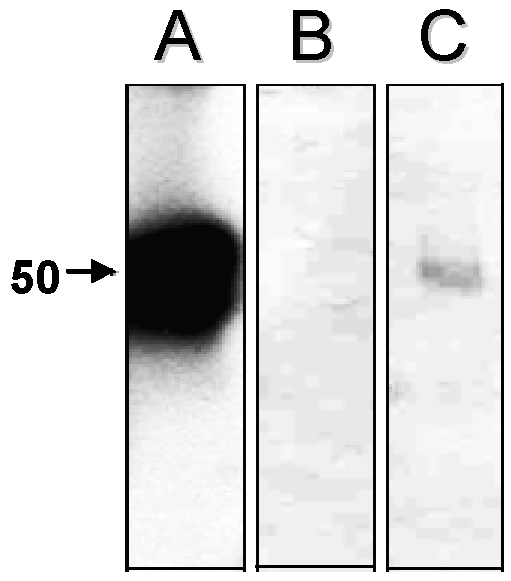
Previous studies showed that cathepsin L is the most active cysteine peptidase in hepatocytes, contributing to more than 40% of their total proteolytic activity, being able to hydrolyze several protein substrates (14). However, we cannot discard the hypothesis that the augment in the cysteine peptidase activity in the T. cruzi-infected hepatocytes was due to expression of the parasite cysteine peptidase named cruzipain, which migrates in gelatin-SDS-PAGE in this same molecular mass range and belongs to the papain C1 family of cysteine peptidases, with a specificity intermediate between those of cathepsin L and cathepsin B (2). Supporting our idea, western blot analysis evidenced a faint polypeptide band with apparent molecular mass of 50 kDa in the T. cruzi-infected hepatocyte cellular extracts after probing with anti-cruzipain polyclonal antibody (Fig. 3, lane C). Conversely, no reactivity was observed when only hepatocyte cells were analyzed (Fig. 3, lane B), demonstrating an absence of immunological cross-reactivity between cysteine peptidases synthesized by the hepatocytes and the parasite.
Corroborating our findings, earlier studies have shown that cruzipain is able to degrade several substrates including gelatin, hemoglobin and casein (15), and this fact may have some positive effect either on the parasite development, penetration and survival inside the host cells (1-3). In previous studies, cruzipain activity has been associated with the growth and differentiation of epimastigotes and amastigotes (2). In order to survive, intracellular organisms must escape the anti-microbial mechanisms of distinct host cells. For instance, cruzipain of T. cruzi participates in the nutrition of the parasite at the expense of the host and is involved in the escape mechanisms of the parasite from the host’s immune system (1, 2). Cruzipain stimulates potent humoral and cellular immune responses during infection in both humans and mice. Furthermore, cruzipain modulates the host response in favor of the parasite survive. Stempin et al. (16) demonstrated that cruzipain decreases NO and IL-12 production from macrophages stimulated by LPS. IL-10, induced by cruzipain in peritoneal cells, is a cytokine that could enhance the alternative activation of macrophages and favor parasite replication. It is possible that IL-10 released by macrophages could deviate the immune system to the T2 response. IL-10 and some microbial antigens are endogenous inhibitors of regulation of IL-12. Then, the balance between anti-inflammatory (IL-10 and TGF-b) and proinflammatory (IL-12) cytokines at the beginning of T. cruzi infection could be crucial for the installation of the parasite (16).
Hepatocyte infections with T. cruzi have been reported in association with bacteria in immunosuppressed mice (17). Soeiro et al. (18) showed that the surface charge of the mouse embryo hepatocytes was affected by T. cruzi, showing a decrease in their Zeta potential of approximately 10% during the first 20 h of parasite-cell interaction. T. cruzi is able to replicate in the liver, suggesting the parasite capability of inhibiting innate immunity. Since most physiological liver activities are due to hepatocyte cells, any disturbance in the number of healthy cells may be pivotal to organ homeostasis and function. For example, during human or experimental visceral leishmaniasis, there is an increase in the production of hepatic transaminases, indicating hepatocyte lesions (19, 20).
In the current study, a cell-associated peptidase of 100 kDa was also detected in both systems (Fig. 1D), which was totally inhibited by 10 mM 1,10-phenanthroline (Fig. 1D, lane Phe), indicating that this should be a metallopeptidase or metal-activated peptidase. The activity of the 100 kDa metallopeptidase was only detected under alkaline conditions and was down regulated in the T. cruzi-infected hepatocytes (Fig. 1D, lane b) when compared with the non-infected ones (Fig. 1D, lane a, and Table 1), showing a clear modulation in the peptidase production along the infective process. When casein and hemoglobin were incorporated into the gel, no proteolytic activity was observed (data not shown)
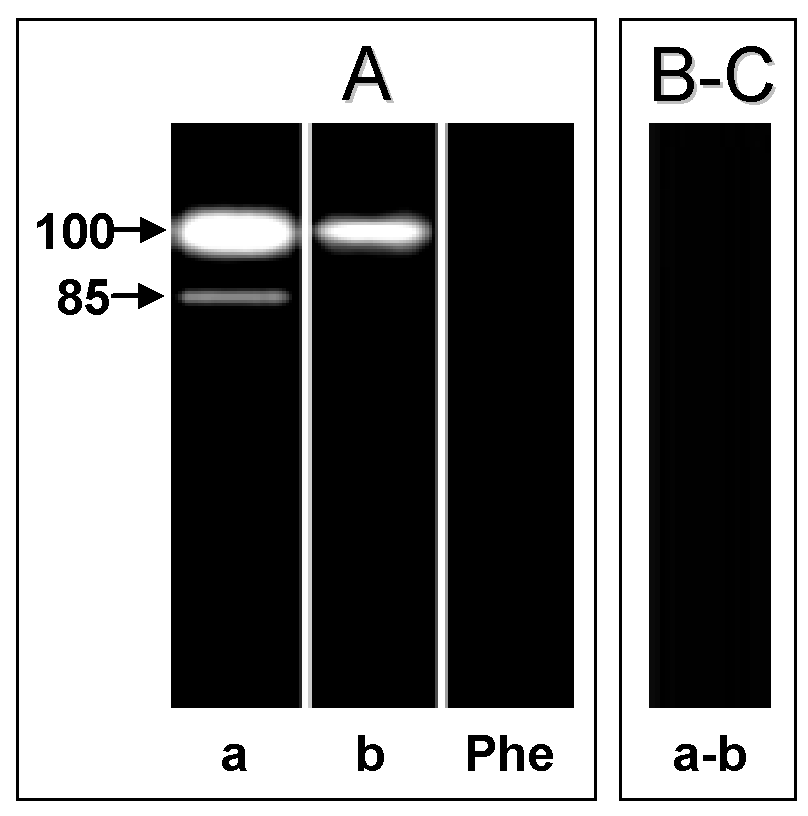
In the cell-free supernatant of both hepatocyte systems, we detected a major metallopeptidase migrating at 100 kDa, active at pH 5.5 (data not shown) and pH 10 (Fig. 4A, lanes a and b). In the non-infected hepatocyte cell-free supernatant we also detected an additional 85 kDa metallopeptidase only at pH 10 (Fig. 4A, lane a), which was also blocked by 10 mM 1,10-phenanthroline (Fig. 4A, lane Phe). Both proteolytic activities were able to degrade only gelatin (Fig. 4B-C). Previous studies have shown by SDS-PAGE the release of gelatin-degrading enzymes migrating at 92 and 95 kDa by human and rat Kupffer cells, respectively (21). These enzymes are named MMP-9 or gelatinase B, and the higher and lower molecular masses of these enzymes correspond to the latent and active forms of MMP-9, respectively (22). Similarly, in a previous work of our research group, we characterized two extracellular metallopeptidases (85 and 100 kDa) as belonging to the MMP-9 family (5). In addition, as visualized in the cellular extract, the 100 kDa extracellular metallopeptidase had its activity significantly reduced in the T. cruzi-infected hepatocytes, while the 85 kDa component was not detected (Table 1).The presence of the parasite in the infected hepatocytes may inhibit a biochemical process that culminates in the secretion of these metallopeptidases into the extracellular environment. The fact that the 85 kDa extracellular metallopeptidase, which corresponds to the active form of MMP-9 (5), has been found exclusively in the supernatant fluid of the non-infected hepatocytes, suggesting a role for this enzyme in the breakdown of the ECM in the hepatocyte cell spreading and growth. It is known that MMPs, including MMP-9, participate in a peptidase cascade to remodel the ECM (23). Earlier studies have described an ECM-degrading peptidase in T. cruzi, and this proteolytic activity may facilitate invasion of host cells, an activity that is likely to be relevant in vivo during the navigation of interstitial tissue spaces by trypomastigote forms (24).
Collectively, the results present herein strongly suggest that expression of cysteine- and metallopeptidase activities is modulated in hepatocyte cells during the T. cruzi infection process. In addition, the fact that T. cruzi is capable of invasion and development of its entire cell cycle within an in vitro system of primary cultures of mouse embryo hepatocytes should make this culture system a useful tool to investigate the pharmacotoxicological effects of new drugs against human Chagas’ disease, since the liver performs numerous important metabolic functions such as detoxification of endogenous and exogenous compounds.
|
|
|
|
|
|
|
ACKNOWLEDGEMENTS |
|---|
This study was supported by Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (MCT/CNPq), Conselho de Ensino para Graduados e Pesquisas (CEPG/UFRJ), Fundação Oswaldo Cruz (FIOCRUZ), Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) and Fundação Universitária José Bonifácio (FUJB). The experiments comply with the current laws of Brazil.
|
|
REFERENCES |
|---|